ANATOMIA Y FISIOLOGIA - Circulación de la sangre
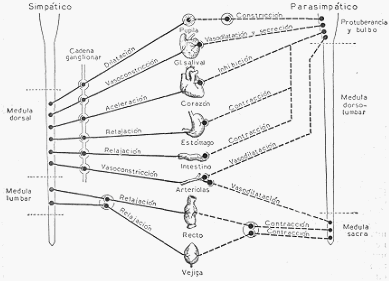
REGULACION NERVIOSA DE LA PRESION ARTERIAL
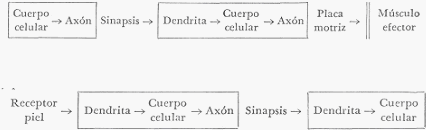
a) Nervios vasomotores. Las arteriolas reciben dos clases de nervios: vasoconstrictores y
vasodilatadores. Los primeros provocan la contracción de los músculos lisos de la pared
arteriolar, y los segundos, su relajación.
Los nervios vasoconstrictores pertenecen a la división ortosimpática del sistema nervioso
autónomo, y los vasodilatadores, a la división parasimpática del mismo sistema.
Los nervios vasoconstrictores actúan constantemente; es decir, que existe un tono
vasoconstrictor que mantiene a las arterias en un estado de permanente semicontracción. La
sección de los nervios vasoconstrictores de un territorio vascular, provoca una vasodilatación
por supresión de dicho tono. Si se excitan entonces los nervios vasodilatadores de esa zona, se
producirá una dilatación aún mayor de los vasos.
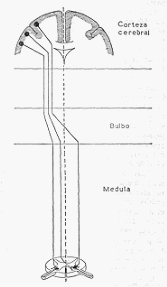
b) El centro vasomotor. La acción tónica continua de los nervios vasoconstrictores, lo mismo
que en el caso del tono del nervio vago, depende de la actividad de un centro vasoconstrictor
situado en el bulbo.
La sección de la medula por debajo del bulbo, produce una caída inmediata de la presión
arterial a niveles bajísimos, por
supresión del tono vasoconstrictor, con la consiguiente
dilatación de las arteriolas de todo el organismo. Una sección del eje cerebroespinal por
encima del bulbo, no modifica el calibre vascular ni afecta la presión arterial.
c) Regulación de la actividad del centro vasomotor. Los factores que mantienen la actividad
tónica del centro vasomotor son de diversa naturaleza. La influencia principal corresponde a
la presencia de anhídrido carbónico en la sangre; al aumentar la tensión del mismo aumenta la
actividad del centro y viceversa. Pero también influyen sobre el centro estímulos nerviosos
provenientes de las regiones superiores del sistema nervioso, o de diversos nervios que llegan
al centro desde la periferia.
Entre los primeros, mencionaremos los efectos de ciertas emociones. El rubor de la vergüenza,
la palidez de ciertos estados emotivos, la congestión de ciertos órganos eréctiles por estímulos
psíquicos, son todos fenómenos vasomotores que tienen su origen en el cerebro. La
vasodilatación puede ser tan generalizada como para provocar una caída pronunciada de la
presión arterial con pérdida del conocimiento. Tal es el origen de algunos desmayos.
Al centro vasomotor llegan también estímulos nerviosos provenientes de los más diversos
orígenes. La estimulación de casi todos los nervios sensitivos produce una elevación de la
presión arterial por aumento del tono vasoconstrictor. En general, cualquier estímulo doloroso
produce vasoconstricción refleja con palidez y elevación de la presión.
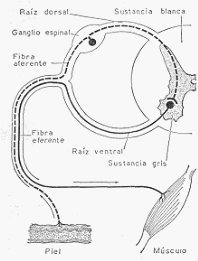
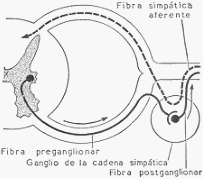
Existen, sin embargo, dos notables excepciones. Los nervios de Hering, que nacen en el seno
carotídeo, y los nervios depresores, que nacen en el cayado aórtico, son nervios sensitivos que
ya hemos descrito al hablar de la regulación de la frecuencia cardiaca. La excitación del cabo
central de estos nervios provoca una acentuada vasodilatación con caída de la presión arterial.
Recordemos que son estimulados normalmente por la presión arterial y que actúan sobre el
centro cardiomoderador provocando bradicardia. Además de esta acción, deprimen al centro
vasoconstrictor.
Como la sangre ejerce continuamente cierta presión sobre las paredes de las carótidas y de la
aorta en las zonas en que nacen los nervios de Hering y depresores, llegan continuamente por
esta vía al centro vasomotor impulsos que frenan su actividad tónica. Si la presión arterial
disminuye por cualquier causa, viajarán menos estímulos por estos nervios, disminuirá el
freno impuesto a la actividad del centro vasomotor, y se producirá una vasoconstricción
generalizada refleja que tiende a corregir la hipa tensión inicial. Si, por el contrario, se produce
un aumento de la presión arterial, llegarán más estímulos al centro vasomotor por la vía de los
nervios depresores y de Hering, el centro disminuirá su actividad tónica, y se producirá un
aumento del calibre arterial que tiende a corregir la hipertensión inicial.
Debido a esta acción frenadora, se ha dado a los nervios de que hablamos el nombre de "los
cuatro frenos de la presión arterial". La sección de estos cuatro frenos produce una
hipertensión arterial permanente con vasoconstricción generalizada y taquicardia.
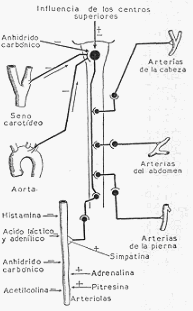
B) INFLUENCIAS HUMORALES SOBRE EL CALIBRE VASCULAR. Aparte de la acción del
anhídrido carbónico sobre el centro vasomotor y de los intermediarios químicos, simpatina y
acetilcolina, a los que se debe en definitiva la acción de los nervios vasomotores sobre el
calibre arterial, existen sustancias formadas en el organismo que son capaces de modificar el
estado de la musculatura vascular.
De estas sustancias formadas en el organismo, unas son productos del metabolismo normal:
anhídrido carbónico, ácido láctico, ácido adenílico, histamina, y otras son productos de
secreción de glándulas de secreción interna: adrenalina, vasopresina, hipertensina.
Centro vasoconstrictor y distribución de los nervios vasoconstrictores.
Los productos del metabolismo de los tejidos tienen, en general, una acción vasodilatadora, la
cual seguramente posee cierta significación fisiológica. En efecto, un músculo en actividad
requiere, como ya lo dijimos, una mayor afluencia de sangre que le aporte el oxígeno necesario
para los procesos químicos que liberan la energía de la contracción. Ahora bien, durante su
actividad el músculo produce anhídrido carbónico, ácido láctico y ácido adenílico, todas
sustancias vasodilatadoras que contribuyen a la dilatación vascular y al mayor aporte
sanguíneo.
Entre los productos de las glándulas de secreción interna, la adrenalina, producto de secreción
de la medula suprarenal, es, sin duda, la de mayor acción. En ciertas condiciones fisiológicas
de "emergencia", como el ejercicio, las emociones violentas, el miedo, y otras, las glándulas
suprarrenales vierten a la circulación un exceso de adrenalina, cuya acción sobre los diversos
sectores del organismo prepara a éste para hacer frente a dichas situaciones. En efecto, la
adrenalina produce vasoconstricción especialmente en el territorio de los órganos del
abdomen y en la piel (ésta es la causa de la palidez), aumento de la frecuencia cardiaca, del
volumenminuto y de la presión arterial, todo lo cual aumenta el aporte sanguíneo a los
músculos.
La vasopresina, producto de secreción del lóbulo posterior de la hipófisis, tiene una acción
vasoconstrictora intensa sobre ciertos territorios vasculares cuando es inyectada; es dudoso,
sin embargo, que desempeñe un papel en la regulación fisiológica del calibre vascular y de la
presión arterial.
La hipertensina es una sustancia que se forma en la sangre cuando el riñón, sometido a una
irrigación sanguínea insuficiente, segrega un producto de secreción interna, la renina. Tiene
acción vasoconstrictora generalizada y es posible que intervenga en la regulación de la presión
arterial en ciertas condiciones extremas.
C) HIPERTENSION ARTERIAL. Una
de
las enfermedades más comunes del hombre, es la
hipertensión arterial. En lugar de las cifras normales de 120 y 80 milímetros de mercurio para
las presiones sistólica y diastólica, se encuentran cifras de 180 a 250 y más, para la sistólica, y
120 a 160 para la diastólica. Los estudios realizados han permitido de' mostrar que en la
hipertensión humana existe un aumento de las resistencias periféricas por vasoconstricción
generalizada. A pesar de las investigaciones realizadas por fisiólogos y médicos de todo el
mundo, todavía no se ha podido averiguar con certeza cuál es el mecanismo causante de este
trastorno.
En los animales se puede provocar experimentalmente una hipertensión arterial permanente
por sección de los "cuatro frenos" (ver más arriba) o por intervenciones sobre el riñón que
limitan su irrigación normal. La hipertensión provocada por este último método, que es la más
parecida a la hipertensión humana, es de origen humoral; es decir, que el sistema nervioso no
es factor determinante en su aparición.
EL SISTEMA LINFATICO
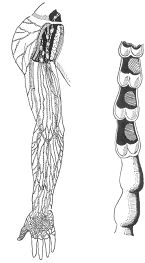
El sistema linfático constituye un anexo del sistema circulatorio.
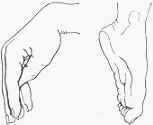
Linfáticos del miembro superior y vaso linfático cortado para mostrar las válvulas.
Los capilares linfáticos, que nacen en los espacios intercelulares y forman troncos que van a
terminar en definitiva en las venas, constituyen, en efecto, una especie de sistema de drenaje
auxiliar del sistema venoso. De la red capilar linfática parten troncos colectores provistos de
válvulas que impiden la vuelta de la linfa a la periferia. Estos colectores van a terminar en el
conducto torácico que recibe la linfa de toda la parte del cuerpo situada por debajo del
diafragma, y de la mitad izquierda de las porciones del cuerpo situadas por encima del
diafragma, siendo el conducto linfático derecho el que recibe los colectores de la mitad derecha
supradiafragmática. El conducto torácico desemboca en la vena subclavia izquierda, y el
conducto linfático derecho en la vena subclavia derecha.
En el trayecto de los conductos linfáticos se intercalan ganglios, constituidos por una red de
tejido conectivo en cuyas mallas se encuentran células de dos tipos: unas que van a convertirse
en linfocitos, uno de los tipos de glóbulos blancos, y otras células (macrófagos) que tienen la
propiedad de englobar toda clase de partículas.
Por los conductos linfáticos circula un líquido de composición diferente según el territorio de
que provenga, y que tiene su origen en los líquidos que bailan las células. Cualquier partícula
o microbio que penetre en un conducto linfático pasará y será retenido en los ganglios
correspondientes. De esta manera, el sistema linfático interviene en los procesos de defensa del
organismo. Todos sabemos que una infección del miembro inferior provoca la tumefacción de
los ganglios de la ingle, así como la de un miembro superior provoca la tumefacción de los
ganglios de la axila.
Los linfáticos provenientes del intestino tienen, además, una intervención preponderante en la
absorción de las grasas. Después de una comida rica en grasas, la linfa del conducto torácico
está constituida por un líquido lechoso que contiene gran cantidad de grasa en forma de gotas
pequeñísimas.
La linfa circula en los vasos linfáticos desde los espacios tisulares hacia las venas en que
desembocan los dos gruesos troncos colectores. Los factores que hacen progresar la linfa son la
acción de masaje y expresión de los músculos y los movimientos respiratorios. También
contribuyen las pulsaciones arteriales, y quizá .las contracciones de las paredes de los propios
vasos linfáticos. Gracias a las válvulas con que éstos están provistos, todas esas fuerzas hacen
progresar la linfa en un solo sentido.
Digestión
Los alimentos que el hombre, lo mismo que otros animales, necesita para poder subsistir, son
de diverso origen: animal, vegetal o mineral. Fuera de estos últimos, constituidos por
sustancias simples, como sales, agua, oxígeno, los alimentos son, en general, sustancias
complejas que el organismo es incapaz de absorber si no sufren un proceso previo de
transformación.
El aparato digestivo está encargado de transformar las sustancias alimenticias complejas
insolubles en sustancias más simples, solubles y difusibles, que pueden ser absorbidas y
utilizadas por el organismo para crecer o reparar sus pérdidas, y para proveerlo de energía. Es
como si se desarmara una casa prefabricada y se utilizaran luego las piezas que la componían
para construir otro edificio, o para reparar una casa que sufre los desgastes producidos por el
tiempo, o para servir de combustible.
ENZIMAS DEL APARATO DIGESTIVO
Los elementos con que cuenta el aparato digestivo para cumplir sus funciones son las enzimas
(fermentos) contenidas en las secreciones de sus glándulas (tabla I). Estas enzimas aceleran la
conversión de las moléculas grandes de proteínas, grasas e hidratos de carbono en sus
elementos constitutivos. Son instrumentos especializados y destinados exclusivamente a la
desmontadura de determinadas partes del edificio. Las enzimas que desarman a las proteínas
hasta reducirlas a aminoácidos, que son sus elementos constitutivos, se denominan proteasas;
pero hay proteasas que sólo son capaces de reducir las proteínas complejas a sustancias
proteicas de menor peso, formadas por grupos de aminoácidos; para completar la digestión
intervienen otras proteasas que atacan a estos bloques más pequeños hasta convertirlos en
aminoácidos.
Las enzimas que transforman a las grasas en ácidos grasos y glicerina son las lipasas. Las que
atacan a los hidratos de carbono se denominan de diferente manera según el sustrato: las que
atacan a los polisacáridos como el almidón se denominan amilasas, y las que atacan a los
disacáridos hasta convertirlos en monosacárido' (como la glucosa) que son absorbidos, son la
sacarasa, maltasa y lactasa.
Además, el aparato digestivo está especialmente adaptado para preparar los alimentos de
manera que puedan ser atacados en la mejor forma por las enzimas. Por su acción mecánica
dividen a los trozos relativamente grandes de alimentos en trozos más pequeños, que se
embeben más fácilmente con los jugos digestivos y, mediante movimientos adecuados, los
hacen progresar desde la boca hasta el ano.
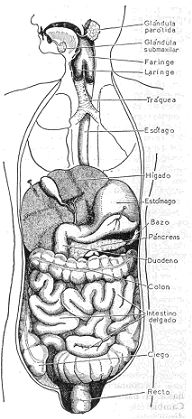
ANATOMIA DEL APARATO DIGESTIVO
Corte vertical del cuerpo humano, para mostrar la distribución y situación exacta de todos los
órganos que componen el aparato digestivo.
A) TUBO DIGESTIVO. En la boca el alimento es masticado y dividido por la acción de los
dientes y, mezclado con la saliva, pasa luego durante la deglución por la faringe y desciende
por
el esófago. Este es un tubo hueco, grueso como el dedo pulgar y de unos 25 cm de
longitud, que se dirige verticalmente hacia abajo, recorriendo la parte inferior y posterior del
cuello y al tórax en toda su extensión, apoyado contra la columna vertebral, para terminar en
el estómago, después de atravesar el diafragma.
El estómago es una víscera hueca situada, por debajo del diafragma y del hígado, en la parte
superior izquierda del abdomen. Su forma suele ser comparada a la de una gaita. Su orificio
superior de entrada, denominado cardias, lo comunica con el esófago, y su orificio inferior de
salida, el píloro, lo comunica con el duodeno. Tanto el cardias como el píloro, están provistos
de fibras musculares circulares (esfínteres) que, por su contracción, cierran el conducto.
Al estómago sigue el intestino delgado, cuya primera porción, que está fijada en la pared
posterior del abdomen, se denomina duodeno (del latín duodeni, que quiere decir doce); tiene
una longitud aproximada de doce dedos. El resto del intestino delgado es el yeyuno-íleon, de
una longitud mucho mayor que el duodeno; se dispone en forma de ovillo y flota en la
cavidad abdominal.
El yeyuno-íleon desemboca en el intestino grueso y su desembocadura está provista de una
válvula: la válvula ileocecal.
El intestino grueso es el segmento terminal del tubo digestivo y forma como una guirnalda
alrededor del intestino delgado. En su origen ocupa la parte inferior y derecha del abdomen;
es el ciego, provisto de una delgada prolongación: el apéndice. Luego se dirige hacia arriba
ocupando el lado derecho hasta llegar debajo del hígado: es el colon ascendente. Allí se acoda,
dirigiéndose horizontalmente hacia la izquierda hasta el bazo: es el colon transverso. Cambia
allí nuevamente de dirección, dirigiéndose hacia abajo (colon descendente) hasta penetrar en
la pelvis (colon ilio pelviano) y terminar en el recto, cuyo orificio de salida o ano está provisto
de anillos musculares, los esfínteres anales.
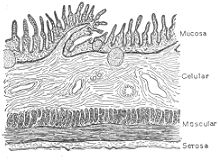
Estructura histológica. En toda su extensión, desde el esófago hasta el recto, el tubo digestivo
presenta una estructura parecida.
Corte longitudinal esquemático del intestino delgado.
Sus paredes están constituidas por cuatro capas concéntricas y superpuestas: 1) una capa
interna, la mucosa, formada principalmente por tejido epitelial y que contiene glándulas que
vierten su secreción en la luz del conducto; 2) una capa celulosa de tejido conectivo ocupada
por vasos y por una
red de nervios y células nerviosas; 3) una capa muscular formada por
fibras circulares y longitudinales dispuestas en distintos planos, y 4) una capa externa formada
por tejido conectivo laxo (capa celulosa) o por el peritoneo, membrana serosa que tapiza las
paredes de la cavidad abdominal y los órganos en ella contenidos.
El peritoneo tiene por función facilitar el deslizamiento de los órganos sobre la pared y los
órganos vecinos, y sostenerlos en posición.
B) GLANDULAS ANEXAS. Además de las glándulas ubicadas en la capa mucosa del tubo
digestivo, existen a lo largo del aparato digestivo varias glándulas anexas que vierten su
contenido en la luz del mismo.
En la boca vierten su secreción tres pares de glándulas salivales: las parótidas, situadas en la
cara por delante del oído, las submaxilares, situadas por debajo de la mandíbula, y las
sublinguales, situadas debajo de la lengua.
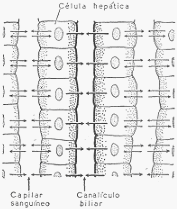
El hígado, la más voluminosa de las vísceras, aparte de sus importantes funciones metabólicas,
segrega la bilis, que llega al duodeno a través del conducto colédoco. El hígado está situado en
la parte superior y derecha del abdomen. Sus células constitutivas se disponen en forma de
lobulillos en los cuales cada célula está en contacto, por uno de sus polos, con un capilar
sanguíneo y, por el otro, con un canalículo biliar. Las células toman del capilar sanguíneo los
materiales con los que elaborarán su doble secreción: por un lado la bilis, que es recogida por
los canalículos biliares y, por otro, productos de secreción interna que vierten en la sangre del
capilar sanguíneo.
Representación esquemática de la disposición de las células hepáticas. Se puede observar el
capilar sanguíneo y el canalículo biliar.
El páncreas es una glándula que vierte su secreción externa en el duodeno. Está situado en la
porción superior del abdomen, por delante de la columna vertebral y por detrás del estómago.
Su porción izquierda o cola toma contacto con el bazo, y su extremidad derecha o cabeza está
rodeada por el duodeno. Las glándulas en racimo que lo constituyen vierten su secreción en
conductos que, después de un trayecto más o menos largo, se unen formando un conducto
colector principal, el conducto de Wirsung. Este se une al colédoco, que trae la bilis del hígado,
para desembocar en el duodeno al nivel de su segunda porción.
CURSO DE LA DIGESTION
La digestión comienza en el momento en que se introducen los alimentos en la boca. Por la
acción combinada de las mandíbulas y de los dientes, de los
músculos masticadores, de la
lengua y de las mejillas, los alimentos son triturados, divididos y mezclados con la saliva hasta
formar un bolo que luego será tragado.
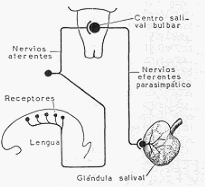
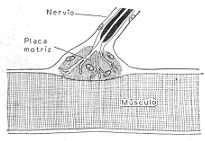
LA SALIVA. Mecanismo de secreción: La saliva es segregada por las glándulas salivales como
consecuencia de un reflejo. Ciertos estímulos térmicos (calor), mecánicos (desecación,
masticación, cuerpos extraños, alimentos) o químicos (ácidos, sustancias amargas, sustancias
alimenticias), excitan terminaciones nerviosas ubicadas en la boca y en la lengua y de donde
parten nervios que llevan sus estímulos a un centro salival ubicado en el bulbo. De allí parten
las fibras simpáticas y parasimpáticas que inervan a las glándulas salivales y provocan su
secreción.
Esquema que representa el mecanismo de la secreción salival. La saliva es segregada como
consecuencia de un reflejo.
La secreción de saliva también puede ser provocada por un reflejo condicionado, cuyo ejemplo
más simple es el de "hacérsenos agua la boca" al ver la comida o pensar en ella. El hecho de
colocar en la boca una sustancia de sabor agradable basta para provocar la secreción de saliva
por la vía refleja que se ha descrito. Si se asocia al acto de proporcionar alimentos un estímulo
cualquiera, incapaz de por sí de provocar secreción salival (p. ej. el sonido de una campanilla),
y se repite un determinado número de veces la aplicación simultánea de ambos estímulos (la
comida y el sonido), llegará un momento en que bastará el sonido de la campanilla para que se
produzca la secreción salival. Se ha adquirido un nuevo reflejo, no innato, que Pavlov, el
famoso fisiólogo ruso, denominó reflejo condicionado.
Función de la saliva. La función de la saliva es múltiple: diluye los alimentos y lubrica el bolo
alimenticio, favoreciendo así la masticación y la deglución; disuelve las sustancias
alimenticias, permitiendo la gustación; humedece las mucosas de la boca, favoreciendo la
fonación (cuando un orador no segrega suficiente saliva se le seca la boca y tiene que recurrir
al clásico sorbo de agua); protege a los dientes e inicia la digestión de ciertos hidratos de
carbono.
La acción digestiva de la saliva se debe a una enzima, la ptialina, amilasa que desdobla el
almidón y el glucógeno —hidratos de carbono de molécula muy grande— en compuestos más
simples hasta llegar a compuestos disacáridos (maltosa). Como la permanencia de los
alimentos en la boca es breve, la ptialina apenas puede ejercer su acción. Sin embargo, el bolo
alimenticio empapado con saliva permanece durante algún tiempo en el estómago antes de ser
penetrado por el jugo gástrico y, durante este tiempo, mientras la reacción del medio no es
demasiado ácida, la ptialina ejerce su acción hidrolítica.
DEGLUCION. Se entiende por deglución el pasaje de los alimentos desde la boca hasta el
estómago. Cuando los alimentos están ya masticados y mezclados con saliva, pasan a un
espacio formado por el hueco del dorso de la lengua y el paladar. La parte anterior de la
lengua se aplica entonces contra el paladar y empuja el bolo hacia la faringe. Una vez llegado
allí, se inician una serie de movimientos reflejos que no dependen de la voluntad y que tienen
corno resultado: 1) cerrar la comunicación de la faringe con las fosas nasales, lo que impide a
los alimentos salir por la nariz; 2) cerrar el orificio superior de la laringe, lo que impide a los
alimentos penetrar en las vías respiratorias. El bolo se desliza entonces penetrando en el
esófago, en donde progresa hacia el estómago gracias a las contracciones de la musculatura
esofágica, que adquieren el carácter de ondas. Estas ondas llamadas peristálticas, de origen
reflejo, y cuya vía eferente es el nervio neumogástrico o vago, consisten en una constricción
localizada en el anillo de las fibras circulares de la pared, constricción que progresa a lo largo
del esófago recorriéndolo en cerca de 5 segundos, y que es precedida por una onda de
relajación.
DIGESTION EN EL ESTOMAGO
A) VACIAMIENTO DEL ESTOMAGO. Apenas llegan los alimentos al estómago se inician en
él, al nivel de la parte media, ondas peristálticas que progresan hacia el píloro.
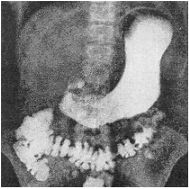
Radiografía del estómago y del intestino grueso, en su posición normal, tomados en conjunto.
Estas contracciones mezclan los alimentos entre ellos y con el jugo gástrico, hasta transformar
el contenido gástrico en una pasta semilíquida que constituye el quimo. Cuando la presión
provocada por las contracciones peristálticas es mayor en el estómago que en el duodeno, pasa
a éste parte del quimo.
El píloro (palabra que deriva de raíces griegas que significan guardián de la puerta) ejerce
efectivamente las funciones de vigilante o portero. Permanece abierto y dispuesto a dejar pasar
cualquier alimento que quiera abandonar el estómago. Si el contenido gástrico es aceptable
para el duodeno, el vaciamiento del estómago se prosigue sin interrupción. Pero si no lo es, se
cierra rápidamente la puerta o píloro y permanece clausurada hasta que una nueva porción
"de prueba" sea aceptada.
Los movimientos del estómago, lo mismo que los del intestino, persisten después de eliminar
todos los nervios que llegan a él. El origen de estos movimientos se halla en la red nerviosa
que ocupa la capa submucosa. Los nervios que llegan a estos órganos proceden del simpático,
cuya excitación provoca relajación muscular, y del parasimpático (vago), cuya excitación
provoca contracción de la capa muscular.
El tiempo de evacuación gástrica es variable según la clase de alimentos ingeridos. El agua y
ciertos líquidos y semilíquidos pasan casi inmediatamente al intestino. Sin embargo, los
líquidos muy ácidos, muy alcalinos o muy concentrados, demoran más en abandonar el
estómago porque al llegar al duodeno lo irritan provocando el cierre del píloro. Los alimentos
sólidos se evacuan tanto más rápidamente cuanto más pronto se convierten en una masa
semilíquida. Los hidratos de carbono se evacuan más rápidamente que las proteínas y éstas
más que las grasas.
B) JUGO GASTRICO. Es el producto de secreción de las glándulas que tapizan la mucosa del
estómago. Las glándulas principales, que ocupan la parte superior del estómago más cerca del
cardias que del píloro, segregan ácido clorhídrico y pepsinógeno. Se denomina así a
una
sustancia proteica que por la acción del ácido clorhídrico se transforma en una proteasa, la
pepsina, capaz de atacar a proteínas de peso molecular elevado, convirtiéndolas en proteosas o
peptonas de peso molecular menor. El ácido clorhídrico también tiene una acción similar, pero
su función principal es la de activar al pepsinógeno transformándolo en pepsina, y
proporcionar a esta enzima el medio ácido necesario para que pueda ejercer su acción.
También segregan las glándulas gástricas el fermento del cuajo, que coagula la leche, y un
factor llamado intrínseco, que participa en la formación de un
principio necesario para la
maduración de los glóbulos rojos de la sangre. Además existe una lipasa que sólo es capaz de
actuar sobre las grasas ya emulsionadas (dispersas en finas gotitas) como las de la yema de
huevo.
La secreción del jugo gástrico se lleva a cabo por dos mecanismos distintos: uno nervioso y
otro humoral.
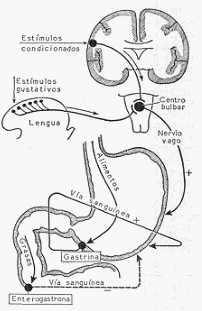
MECANISMO DE SECRECION DEL JUGO GASTRICO. La excitación del vago provoca la
secreción del jugo gástrico. La presencia de comida en la boca provoca una sensación gustativa
que por vía refleja, y a través de los nervios vagos, determina una secreción gástrica. Basta sin
embargo pensar en la comida, verla u olerla, para que se provoque lo que Pavlov denominó
secreción psíquica o de apetito. Este es un reflejo condicionado adquirido por la asociación
frecuente de dos estímulos, uno capaz y otro incapaz de por sí de provocar secreción gástrica
(ver secreción de saliva). La sensación de apetito y la secreción refleja de origen gustativo,
constituyen la llamada fase cefálica de la secreción gástrica.
La llegada de los alimentos al estómago puede constituir, por la distensión que provoca, un
nuevo estímulo para la secreción gástrica. Pero aquí interviene un mecanismo humoral mucho
más importante. Ciertos alimentos, en especial productos de la digestión parcial de las
proteínas, provocan la liberación, por la mucosa del píloro, de una sustancia, la gastrina, que
es absorbida en la sangre, y por esta vía llega a las glándulas principales del estómago, a las
que excita químicamente, provocando su secreción. Esta etapa es la llamada fase gástrica de la
secreción del estómago.
Al pasar los alimentos al duodeno y yeyuno, la secreción gástrica vuelve a ser influida por un
mecanismo humoral. Algunos productos de la digestión, al ser absorbidos y llegar por vía
sanguínea a las glándulas gástricas, estimulan su secreción. Por otra parte, la llegada de grasas
al intestino provoca la libe radón, por la mucosa del duodeno, de una sustancia denominada
enterogastrona, que es absorbida en la sangre y por esta vía llega a las glándulas gástricas
inhibiendo su secreción.
Representación esquemática del mecanismo por medio del cual se produce la secreción del
jugo gástrico.
DIGESTION EN EL INTESTINO
A) MOVIMIENTOS DEL INTESTINO DELGADO.
Al salir del estómago los alimentos se
hallan ya constituyendo una pasta semilíquida, el quimo. El quimo atraviesa rápidamente el
duodeno y recorre el intestino delgado gracias a los movimientos peristálticos del mismo.
Además de los movimientos peristálticos propulsores, aparecen en el intestino delgado, a
intervalos regulares, contracciones que lo dividen, como a una salchicha, en pequeños
segmentos. Estas contracciones no hacen progresar el contenido intestinal; pero, en cambio,
favorecen la mezcla del contenido intestinal con los jugos digestivos y aumentan el contacto de
dicho contenido con las vellosidades de la mucosa intestinal, lo que favorece la absorción.
En el duodeno se mezcla el quimo con las secreciones del hígado y del páncreas.
B) LA. BILIS. La bilis segregada por las células hepáticas pasa de los canalículos biliares a
conductos excretores que se reúnen en un tronco común: el conducto hepático, que se continúa
con el colédoco.
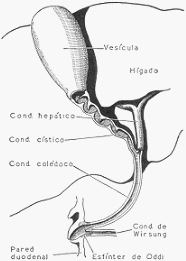
Vías de excreción de la bilis. A partir de la vesícula se observa, en corte longitudinal, el
conducto cístico, el colédoco, el de Wirsung, así como la pared duodenal y esfínter de Oddi.
En éste desemboca el conducto cístico, que lo comunica con la vesícula biliar, reservorio en el
cual se acumula, concentra y modifica la bilis.
El hígado produce constantemente bilis que, en los períodos interdigestivos, pasa a la vesícula,
donde se concentra por absorción de agua y sales y se enriquece con el mucus segregado por
las células de la mucosa de este reservorio. El colédoco está provisto al nivel de su
desembocadura en el duodeno de un esfínter (esfínter de Oddi), cuya contracción impide la
evacuación de la bilis.
La llegada de ciertas sustancias al duodeno (salinas, ácidas, grasas) provoca una relajación del
esfínter de Oddi de probable origen reflejo nervioso y la consecuente entrada de bilis en el
duodeno. Contribuye a esto la contracción de la vesícula biliar, estimulada por vía humoral
por una sustancia denominada colecistoquinina y que se forma en la mucosa del duodeno por
la acción de los productos alimenticios indicados, y que es luego vertida en la sangre.
La bilis contiene, además de mucina, sales biliares, pigmentos biliares, lecitina y colesterol. Su
acción digestiva se debe principalmente, como veremos luego, a las sales biliares.
C) EL JUGO PANCREATICO. En ayunas, la secreción de jugo pancreático se interrumpe por
completo. Pero, como observó Claudio Bernard por primera vez, la ingestión de alimentos
provoca, a los pocos minutos, una abundante secreción que dura un par de horas.
La secreción de jugo pancreático obedece a dos mecanismos: uno nervioso y otro humoral.
Al llegar los alimentos a la boca inician un reflejo que por vía del nervio vago estimula a la
secreción pancreática.
Pero el principal estímulo de la secreción pancreática es el que le llega por la sangre.
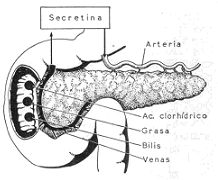
Mecanismo de la formación de la secretina. La secretina se forma en la pared duodenal, es
absorbida por la sangre y llega al páncreas por su arteria nutricia.
La llegada de ácidos, grasas y bilis al duodeno, provoca en la mucosa duodenal la formación
de una sustancia, la secretina, que, absorbida por la sangre, llega al páncreas por sus arterias
nutricias y estimula poderosamente la secreción de sus glándulas.
El jugo pancreático es rico en enzimas; contiene una poderosa proteasa, la tripsina, que es en
realidad una mezcla de enzimas. Lo mismo que la pepsina, la tripsina se segrega en forma de
proenzima, el tripsinógeno, que es activada por un componente del jugo intestinal
denominado enteroquinasa, que lo convierte en tripsina. Contiene también una enzima que
coagula la leche, una poderosa esterasa que hidroliza las grasas, una amilasa y una maltasa.
D) EL JUGO ENTERICO. Se denomina jugo entérico o intestinal al producto de secreción de
las glándulas de la mucosa del intestino delgado. Poco se sabe todavía de los mecanismos que
regulan su secreción. Además de la enteroquinasa ya descrita, contiene proteasas (erepsina),
una lipasa y varias carbohidrasas.
TRANSFORMACIONES DEL QUIMO EN EL INTESTINO DELGADO
La acción de la bilis, del jugo pancreático y del jugo entérico terminan la digestión de las
proteínas, grasas e hidratos de carbono, hasta convertirlos en sus elementos constitutivos que
son luego absorbidos.
Las proteínas, que ya habían sufrido un primer ataque por la pepsina del estómago, son
atacadas por la tripsina y la erepsina hasta convertirlas en aminoácidos que se absorben a
través de la mucosa intestinal.
Las grasas sufren en el estómago su primera transformación por obra de la lipasa gástrica si es
que llegan emulsionadas, es decir, divididas en finísimas gotas. Luego, la lipasa pancreática
actúa sobre el duodeno junto con la bilis. La lipasa pancreática actúa sobre las grasas tanto
mejor cuanto más emulsionadas se hallen éstas. La bilis o, más precisamente, las sales biliares,
desempeñan un papel muy importante en este sentido. Disminuyen la tensión superficial en el
límite aguagrasa y, de esta manera, provocan la división de las grasas en pequeñas gotas, lo
que facilita la acción de la lipasa que las desdoblará en sus componentes: glicerina y ácidos
grasos. Además, los ácidos biliares facilitan la absorción de los ácidos grasos liberados en la
digestión de las grasas.
Los hidratos de carbono requieren todavía más etapas para su total digestión. La digestión del
almidón y el glucógeno —moléculas muy grandes— comienza en la boca por la acción de la
ptialina de la saliva, y continúa en el estómago, mientras la reacción del medio no sea
demasiado ácida, por la acción de la misma enzima. El producto final de la digestión por la
ptialina no pasa del estado de disacárido. La amilasa del páncreas es capaz de atacar también
los azúcares complejos como el almidón, intacto o parcialmente hidrolizado; pero la digestión
de estos azúcares complejos llega en el intestino hasta el estado de azúcares simples
(monosacáridos) por la acción de la maltasa del jugo pancreático y del jugo intestinal, enzima
que desdobla la maltosa en dos moléculas de glucosa, de la invertasa y la lactasa del jugo
intestinal. La primera desdobla la sacarosa en fructosa y glucosa; la segunda convierte a la
lactosa en glucosa y galactosa.
DIGESTION EN EL INTESTINO GRUESO
Aproximadamente a las 4 6 5 horas de haber comenzado la evacuación gástrica comienza a
entrar en el ciego el contenido del intestino delgado, que es una masa semilíquida constituida
principalmente por algunos restos de secreciones (jugo gástrico, pancreático o intestinal,
mucus, bilis), células epiteliales, mucus y leucocitos.
A) MATERIAS FECALES. En el intestino grueso se absorbe una buena cantidad de agua y se
añaden algunos elementos sólidos entre los cuales los más importantes son las bacterias. Se
forma así una pasta semisólida que ha recibido el nombre de heces o materias fecales.
En el intestino grueso existe una enorme cantidad de microorganismos. En un miligramo de
materias fecales existirían, según Cohendy, 144 millones de gérmenes. Los microbios del
intestino grueso pueden clasificarse en dos grupos según el alimento que utilizan: los que
viven de sustancias proteicas provocan su putrefacción, para lo cual no necesitan oxígeno por
ser microorganismos anaerobios. El olor característico a putrefacción está dado por los
productos de la actividad de estos microbios: aminas, ácidos volátiles, fenoles, cuerpos
sulfurados y gases.
El otro grupo de microbios vive de los azúcares, a los que transforman por fermentación en
cuerpos más simples: ácidos, alcoholes, acetonas y gases.
Investigaciones recientes han demostrado que los microbios intestinales pueden sintetizar
algunas vitaminas que luego son absorbidas y utilizadas por el organismo.
B) MOTILIDAD DEL INTESTINO GRUESO. La motilidad del intestino grueso es escasa. Los
movimientos peristálticos parciales sirven para mezclar el contenido del colon y favorecer la
absorción de agua. Sin embargo, pocas veces al día, ya sea a raíz de una comida (reflejo
gastrocólico), de una defecación o de algún otro estímulo reflejo, se inicia en el colon
transverso una contracción intensa que se propaga en forma de onda hacia el colon
descendente, empujando en masa hacia el colon pelviano todo el contenido de la porción distal
del intestino grueso. En el colon pelviano permanecen las materias fecales hasta el momento
de la defecación.
LA DEFECACION
Cuando las materias fecales llegan al recto y lo distienden, se inicia un reflejo que provoca una
contracción peristáltica del intestino grueso y la dilatación de los esfínteres del ano, de lo que
resulta el vaciamiento de todo el intestino grueso a partir del colon transverso. Ayuda a la
evacuación la contracción de los músculos del abdomen, combinada con la contracción del
diafragma con cierre de la glotis.
Riñón
En el incesante flujo de sustancias nutritivas a través del organismo, y los constantes cambios
que éstas sufren con el fin de proveer al organismo de sustancias energéticas o plásticas, se
acumulan productos de desecho que deben ser eliminados. El pulmón excreta anhídrido
carbónico y agua; la piel, agua y algunas sales; por el intestino se eliminan algunos metales
pesados y residuos de los alimentos ingeridos y de las secreciones digestivas; el hígado excreta
los productos finales de la destrucción de hemoglobina, etc. Todas estas excreciones
contribuyen a mantener constante el "medio interior", constancia que es condición
indispensable para la vida independiente del organismo. La importancia del riñón en este
sentido es preponderante, no sólo por la cantidad, sino también por la diversidad de
productos que se eliminan con la orina.
ANATOMIA
Los riñones están situados en el abdomen, por debajo del diafragma, uno a cada lado de la
columna vertebral.
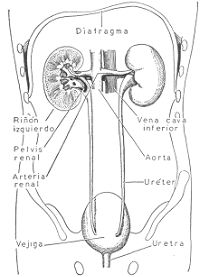
Representación semiesquemática de los riñones y de las vías de excreción de orina. En esta
vista posterior se puede observar el riñón izquierdo cortado perpendicularmente, y la
inserción de la arteria renal y vena cava inferior en los riñones.
En el borde interno de cada riñón existe una escotadura o hilio por donde penetra la arteria
renal, rama de la aorta que pronto se ramifica en la masa del riñón, y por donde sale la vena
renal que desemboca en la vena cava inferior. En la pelvis renal desembocan los tubos
colectores renales que conducen la orina. La pelvis se continúa con el uréter, conducto largo y
del calibre de un lápiz que desemboca en la vejiga. Las paredes del uréter están provistas de
fibras musculares lisas cuyas contracciones peristálticas hacen progresar la orina hacia la
vejiga. Es ésta un órgano hueco que permite la acumulación de la orina y que la evacua al
exterior a través de la uretra. La pared de la vejiga está provista de una túnica muscular
formada por fibras musculares lisas que se disponen en tres capas concéntricas. La capa
media, constituida por fibras circulares, forma alrededor del orificio uretral un músculo
anular, el esfínter interno, cuya contracción, independiente de la voluntad, asegura la
retención de orina en la vejiga. Está reforzado este esfínter por fibras estriadas dispuestas en
forma circular alrededor de la primera porción de la uretra, constituyendo el esfínter externo,
cuya contracción voluntaria refuerza la acción continente del esfínter interno.
MICCION
La orina que penetra en la vejiga impulsada por las contracciones peristálticas del uréter, la
llena en forma lenta. La vejiga se dilata progresivamente, adaptando su capacidad a los
cambios de su contenido sin variar mucho su presión. Cuando pasa de cerca de medio litro la
cantidad de orina contenida en la vejiga, la presión intravesical aumenta considerablemente y
aparecen contracciones rítmicas de musculatura, acompañadas de sensaciones dolorosas; es lo
que constituye el deseo de orinar. La micción puede todavía evitarse voluntariamente
mediante la contracción del esfínter externo.
El acto de la micción es de naturaleza refleja, pero en su iniciación interviene, por lo menos en
los adultos sanos, la voluntad. En los niños y en ciertos casos patológicos es un acto puramente
reflejo. Cuando sobreviene el deseo de orinar en el adulto, éste interviene con su voluntad, ya.
para reprimir el deseo, ya para ceder a él. En el primer caso contrae voluntariamente el esfínter
externo, lo que trae por vía refleja una relajación de la musculatura de las paredes vesicales. En
el segundo caso suprime la contracción voluntaria del esfínter externo, pasa una pequeña
cantidad de orina a la uretra y se inicia una serie de reflejos que determinan la relajación del
esfínter interno y la contracción de la musculatura vesical, vaciándose la vejiga con fuerza.
Para apoyar la fuerza propulsora de la contracción vesical el sujeto hace una inspiración
profunda, cierra la glotis para evitar la salida del aire inspirado y contrae los músculos del
abdomen. Al fin de la micción se contrae otro músculo que rodea la uretra y que contribuye a
expulsar las últimas gotas de orina contenidas en este conducto. En la mujer no existe este
músculo y la micción termina bruscamente.
ESTRUCTURA DEL RIÑON
En un corte de riñón que siga su eje longitudinal, se observa que está formado por dos zonas
diferenciables a simple vista: una porción exterior, la corteza, y una interior, la medula. Un
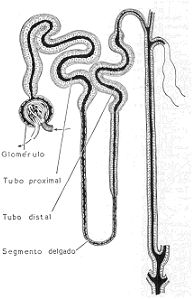
examen microscópico revelará que el riñón está constituido por millares de unidades (los
nefrones) compuestas por tubos en cuyo extremo ciego se invagina un racimo de vasos
capilares sanguíneos (glomérulo), y que después de un trayecto tortuoso y largo forman tubos
colectores ubica dos en la medula, que se van uniendo entre sí hasta vaciarse por fin en la
pelvis renal.
Representación esquemática del nefrón. Obsérvese que el glomérulo está formado por un
racimo de vasos capilares, que resultan de la división de la arteriola aferente (más gruesa) y
que se resumen en la arteriola eferente (más delgada).
La parte del tubo que rodea al glomérulo (cápsula de Bowman) está revestida por células
epiteliales planas y delgadas. En el resto del tubo urinífero se distinguen tres segmentos: 1) el
segmento proximal, provisto de células piramidales recubiertas con un ribete en cepillo; 2) el
segmento delgado, que forma un asa, provisto de células aplanadas, y 3) el segmento distal,
con células piramidales menos altas que las del segmento proximal y sin ribete en cepillo.
La arteria renal, después de llegar al hilio se divide y subdivide, y da por fin ramas que
ocupan la zona limitante entre la corteza y la medula. De allí parten arterias que penetran en la
corteza y de las que emergen las arteriolas aferentes, que, al penetrar en el extremo ciego del
tubo renal, forman una red capilar, el glomérulo. Después de atravesar el glomérulo, la sangre
pasa a una arteriola eferente que se divide pronto en una red capilar que rodea al resto del
tubo urinífero.
MECANISMO DE LA FORMACION DE ORINA
Todos los elementos componentes de la orina, aparte de algunas sustancias como el ácido
hipúrico y el amoníaco, que el riñón es capaz de sintetizar, provienen de la sangre.
Al nivel del glomérulo los capilares sanguíneos están en contacto íntimo con las paredes de la
cápsula de Bowman. Las paredes del extremo ciego del tubo renal están tapizadas por células
chatas muy delgadas que se asemejan a las paredes de los capilares. A través de esta doble
pared filtra hacia el interior del tubo renal un líquido cuya composición, excepto la falta de
proteínas, es idéntica a la del plasma sanguíneo. Las moléculas tienen un tamaño demasiado
grande y no pasan a través de los pequeños poros del filtro glomerular. La fuerza que provoca
la filtración es la presión de la sangre en los capilares sanguíneos que constituyen el
glomérulo. En estos capilares la presión sanguínea es mayor que en los capilares del resto del
cuerpo, principalmente porque el glomérulo está intercalado entre dos arteriolas de las cuales
la eferente es de menor calibre que la aferente. En lugar de 30 mm de mercurio que es la
presión habitual en los capilares de la piel, la presión sanguínea de los del glomérulo se eleva a
70 mm de mercurio. Esto asegura la filtración de un volumen elevado de líquido, que en el
hombre totaliza unos 125 cm3 por minuto, es decir, cerca de 170 litros por día.
La composición de este líquido, como ya dijimos, es idéntica a la del plasma, salvo que no
contiene proteínas. Richards y sus colaboradores han podido probar esto, en los animales de
experimentación, aspirando con micropipetas el contenido de la primera porción del tubo
renal y sometiéndolo a análisis químico.
Si comparamos la composición de la orina con la del plasma (tabla II) veremos, sin embargo,
que la concentración en que se encuentran sus componentes es muy variable, y que algunas
sustancias que existen en la sangre no están presentes en la orina. Esto ocurre porque el
infiltrado glomerular es reabsorbido parcialmente a lo largo del tubo urinífero, y esta
absorción es selectiva.
Consideremos en primer lugar la reabsorción de agua. Mencionamos antes que se filtran a
través de los glomérulos de ambos riñones unos 170 litros de líquido por día. Todos sabemos
que el volumen de orina evacuado por la vejiga es de cerca de un litro y medio. Se deduce de
esto que al nivel de los tubos renales se reabsorben en las 24 horas 168 litros y medio, es decir
el 99% del filtrado glomerular. Intervienen en este proceso solamente los tres segmentos
descritos del tubo urinífero. Los tubos colectores, el uréter, la vejiga, etc., no tienen funciones
de absorción.
Los túbulos reabsorben agua y las sustancias disueltas en ella. Pero la reabsorción de las
sustancias disueltas se hace en forma selectiva. Un ejemplo típico es la glucosa. Esta se
reabsorbe íntegra en el segmento proximal del tubo renal. La capacidad de reabsorción de las
células tubulares es muy grande, pero no infinita. En efecto, cuando la concentración de
glucosa en la sangre, y por consiguiente en el filtrado glomerular, se eleva (como en el caso de
la diabetes), las células del tubo proximal reabsorben más glucosa. Pero pasado cierto límite
denominado umbral renal, que para la glucosa es de alrededor de 1,5 g por litro de sangre, los
túbulos son incapaces de reabsorber toda la glucosa que contiene el filtrado glomerular,
dejando pasar cierta cantidad a la orina. Aparece entonces glucosuria, uno de los síntomas de
la diabetes.
Otra prueba de la reabsorción selectiva lo constituye el grado diferente de concentración que
sufren las sustancias al pasar de la sangre a la orina. Algunos constituyentes se concentran
mucho, mientras otros casi no se concentran (ver tabla II). La urea se concentra 80 veces,
mientras que el cloruro de sodio sólo se concentra 2 veces. Si la reabsorción de agua fuera
pasiva, todas las sustancias contenidas en el filtrado glomerular tendrían que •presentar un
índice de concentración semejante.
También se ha probado por diversos métodos que las células de los tubos uriníferos, así como
son capaces de tomar de la luz de los tubos sustancias que luego vierten en la sangre, son
capaces también de verter en la luz de los tubos sustancias que han tomado de la sangre. El
papel que esta función excretora puede tener en la formación de orina, no está aún aclarado.
OTRAS FUNCIONES DEL RIÑON
Aparte de su función eliminadora de residuos del metabolismo y de sustancias tóxicas, el
riñón desempeña otras funciones igualmente importantes.
A) Regula el volumen del plasma sanguíneo y del agua del organismo. A una ingestión
abundante de agua, el riñón responde eliminando una gran cantidad de orina muy diluida; a
una pérdida abundante de agua, como sucede cuando se suda profusamente, o a consecuencia
de vómitos, diarrea, etc., responde eliminando una cantidad pequeña de orina concentrada.
Esta función reguladora está bajo la dependencia de una hormona segregada por el lóbulo
posterior de la hipófisis, la hormona antidiurética, que actúa sobre las células tubulares
acelerando la reabsorción de agua. Cuando el organismo está deshidratado, la concentración
de hormona en la sangre aumenta y las células tubulares reabsorben más agua. El agua vuelve
al organismo y la orina se concentra. Cuando el organismo está muy hidratado, disminuye la
concentración de la hormona antidiurética en la sangre y las células tubulares reabsorben
menos agua, resultando una orina más diluida y de volumen aumentado. La ausencia de
hormona del lóbulo posterior de la hipófisis provoca una enfermedad llamada diabetes
insípida, que hace eliminar cantidades enormes de orina, conociéndose el caso sorprendente
de un paciente que orinaba 43 litros por día.
B) Regula el equilibrio de ciertos elementos inorgánicos que entran en la composición de la
sangre, mediante la excreción, en mayor o menor cantidad, de iones tales como sodio, potasio,
calcio, cloruros, fosfatos, etc.
B) Regula la reacción de la sangre y, por consiguiente, del organismo. Si en el curso del
metabolismo aumenta la producción de sustancias ácidas, el riñón elimina una orina más
ácida y fabrica amoníaco para ahorrarle bases al organismo. Si aumenta la producción de
bases, el riñón elimina una orina más alcalina y ahorra ácidos.
Las glándulas endocrinas
Las glándulas de secreción interna
o endocrinas son varias: la hipófisis,
el páncreas, las
suprarrenales, las sexuales, etc., y cada una tiene estructura y función propias. Aunque
conocida su estructura anatómica desde mucho tiempo antes, sólo entre 1869 y 1889 el
investigador francés Brown
Sequard estableció el concepto actual de glándula de secreción
interna.
CONCEPTO ACTUAL
Se consideran glándulas de secreción interna las que elaboran productos activos, específicos de
cada una, que vierten en la sangre, regulando a distancia determinadas funciones.
Hay en el organismo glándulas que elaboran productos específicos, pero que los vierten al
exterior y no en la sangre, como ocurre con las salivales y demás glándulas digestivas,
llamadas glándulas de secreción externa. A estos efectos, el tubo digestivo se considera como
si se hallara en el exterior. Se llaman mixtas las que, como el páncreas, tienen una secreción
externa (jugo pancreático) y otra interna (insulina).
La importancia de estas glándulas deriva de que son indispensables para el mantenimiento de
la especie y del individuo normal. En efecto, gobiernan todos los actos de la reproducción, del
embarazo y luego de la lactancia, en forma tal que sin ellas nada de esto es posible. En cuanto
al individuo, presiden su crecimiento y su nutrición, regulándolos, y contribuyen de manera
preponderante al mantenimiento de la unidad orgánica, por coordinaciones múltiples que
establecen entre sus partes.
REGULACION DE SU FUNCIONAMIENTO
Las glándulas de secreción interna elaboran sus productos y los vierten en la sangre, en
cantidad adecuada a las necesidades del organismo. Esta regulación, que suele ser muy
delicada, se efectúa por medio de dos mecanismos, uno nervioso y el otro humoral.
A) MECANISMO NERVIOSO. El mecanismo nervioso es más o menos importante, según la
glándula de que se trate. Así, la secreción de adrenalina por la medula suprarrenal depende
fundamentalmente de estímulos que le llegan por vía nerviosa; basta excitar sus nervios con
una corriente eléctrica para que aumente, y por su sección casi desaparece. En cambio, en el
páncreas endocrino pasa lo contrario: con nervios o sin nervios, la secreción de insulina es
siempre casi normal.
Llama la atención, si se las observa con un microscopio, que en todas las glándulas de
secreción interna existe una riquísima red nerviosa, en comunicación, por un lado, con las
células endocrinas, y por el otro, con el resto del sistema nervioso. El papel que desempeña y
los estímulos que la recorren, son conocidos sólo parcialmente.
B) MECANISMO HUMORAL O SANGUINEO.
El mecanismo humoral de regulación es de
gran importancia y se lo puede clasificar en dos grupos, según los estímulos provengan o no
de otra glándula endocrina.
Cada glándula endocrina fabrica sustancias que vierte en la sangre y llegan a las otras
glándulas endocrinas, a las que estimulan o inhiben, y a su vez, por igual mecanismo, la
primera es influida por las otras. Esto crea un complejo y estrecho sistema humoral de
vinculación entre todas las glándulas endocrinas. En este sentido corresponde a la hipófisis el
lugar preponderante. De esta glándula parten sustancias que por vía sanguínea alcanzan la
tiroides, la suprarrenal, etc., y regulan su forma y su actividad; a su vez, de estas glándulas
parten otras sustancias que, también por la sangre, regulan a la hipófisis.
Este mecanismo de vinculación es de tal importancia que basta provocar una alteración en uno
de los puntos del complicado sistema para que se desarregle del todo, como la máquina de un
reloj cuando se le toca una pieza. Esta sinergia funcional explica claramente por qué son tan
difíciles de estudiar la fisiología o las enfermedades de una glándula de secreción interna.
El segundo grupo de hechos del mecanismo humoral de regulación, comprende los casos en
los cuales existe en la sangre una sustancia química, que actúa de manera específica como
reguladora de una glándula de secreción interna. Esto es lo que sucede con el contenido de
glucosa en la sangre arterial que entra al páncreas: si es grande, aumenta la descarga de
insulina en la sangre venosa, y si es pequeña, disminuye la respuesta.
En resumen, la producción de sustancias específicas por una glándula endocrina y su descarga
ulterior en la sangre, son el resultado complejo de un conjunto de estímulos que recibe por los
nervios o por la sangre, haciéndola de esta manera solidaria con el resto del organismo.
HORMONAS. ACCIONES FISIOLOGICAS
El producto específico fabricado por las glándulas endocrinas y luego vertido en la sangre se
llama hormona, y cada glándula fabrica una o más. Las hormonas tienen el papel de
"mensajeros químicos", porque transportadas por la sangre actúan a distancia de su sitio de
origen, sobre determinados receptores. Las hormonas y el sistema nervioso son los dos
grandes mecanismos de correlación, entre las partes distantes del organismo, en que se basa la
unidad de los seres superiores, constituidos por un conjunto de células de distinta forma y
función. Sistema nervioso y glándulas endocrinas representan juntos lo que para un país su
gobierno central.
Las hormonas pasan a la sangre en cantidades minúsculas, a veces muy difícil o hasta
imposible de descubrir por los más sensibles métodos de análisis de que se dispone. Este
pasaje es continuo y sufre aumentos y disminuciones en diversas circunstancias fisiológicas.
La acción de las hormonas consiste en regular funciones determinadas. Así, algunas de ellas
hacen que el crecimiento del sujeto sea normal, y cuando faltan o están en exceso, éste no sólo
se altera en su talla, sino que es disarmónico; otras son necesarias para que la psiquis se
desarrolle normalmente, y en este caso las alteraciones de ciertas hormonas llevan a trastornos
mentales variables de idiocia o. excitabilidad; las hay que son indispensables para las
funciones de reproducción, las de nutrición y otras tan importantes que sus alteraciones llevan
a la muerte.
Un hecho notable y de gran importancia práctica es que las hormonas, por lo general, no
tienen especificidad zoológica, es decir. que la misma hormona es activa en numerosos
animales de la escala animal. Un ejemplo es la insulina extraída del páncreas de oveja, cerdo o
vaca, que es activa en cualquiera de estas especies y también en el hombre.
El progreso realizado por la endocrinología en sus pocos años de existencia es enorme y
continuamente aparecen nuevos adelantos. Con todo, quedan grandes problemas por resolver.
Por ejemplo, no se conoce en muchas glándulas el número de hormonas que producen, y ni
siquiera se sabe la composición química exacta de algunas o su modo íntimo de actuar.
METODOS DE ESTUDIO
Como son órganos de función tan compleja, su estudio ha sido abordado desde diversos
ángulos. Los primeros estudios fueron de orden anatomoclínico, es decir, se observaba en los
pacientes la evolución de la enfermedad y luego, en la autopsia, se trataba de investigar las
causas. Ha sido útil este camino, pues ha permitido establecer, por ejemplo, que la enfermedad
de Addison era producida siempre por lesión de las suprarenales. En cambio, en otros casos
este método no da informaciones útiles.
Los métodos experimentales, de gran desarrollo en este siglo, han significado
un eficaz
progreso en este terreno, y a ellos se deben casi todos nuestros conocimientos actuales. Se los
aplica en formas diversas. Unas veces en el estudio de las modificaciones anatómicas de las
glándulas, bajo la influencia de un factor dado; otras en la aplicación de técnicas químicas,
tratando de aislar y conocer los productos fabricados por las glándulas, su composición, etc.; y
finalmente, en la producción de alteraciones de una glándula en los animales de laboratorio,
para estudiar los síntomas que aparecen luego. Este último método o fisiológico, es
ampliamente utilizado y permite producir hipofunciones o hiperfunciones en cada una de las
glándulas. Un ejemplo es la castración, que da síntomas de hipofunción genital; y, por el
contrario, el injerto o la inyección de extractos en animales normales, pueden dar los síntomas
de hiperfunción.
Hipófisis
La hipófisis es considerada como la glándula de secreción interna de mayor valor, tanto para
el individuo como para la especie animal a que pertenece. En efecto, sus trastornos perturban
de manera grave casi todas las funciones orgánicas (crecimiento, urinaria, nutritivas, etc.) y las
de reproducción.
Con respecto a las restantes glándulas endocrinas tiene una posición central, de comando,
expresada en diversas formas literarias como cuando se la designa "centro de la constelación
endocrina", "director de la orquesta endocrina", etc.
En el hombre tiene la forma de una judía de tamaño mediano, acostada transversalmente en el
medio y dentro de la base del cráneo, en un receptáculo óseo llamado "silla turca"; su peso es
de 0,5 a 0,7 gramos. Por encima de ella, y en comunicación directa por un corto tallo, se halla el
hipotálamo, que es una importante región del cerebro situada en la parte inferior y media del
mismo.
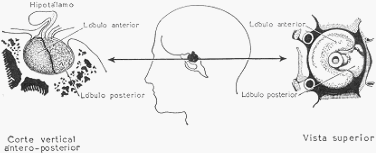
Esquema que representa un corte vertical antero posterior y una vista superior de la hipófisis,
así como su ubicación en la silla turca, en la base del cráneo
El estudio cuidadoso revela que la hipófisis se compone de 3 partes, que son, yendo de
adelante atrás: el lóbulo anterior o pars glandularis; la parte intermedia; y el lóbulo posterior o
pars nervosa.
Al examen microscópico cada una de las tres partes muestra una estructura característica. El
lóbulo anterior está formado por cordones celulares enrollados como una madeja de hilo, entre
los cuales corren capilares sanguíneos. Las células son de 3 tipos en su aspecto y en su función:
sin gránulos en el protoplasma (cromófobas); con gránulos que se tiñen con colorantes ácidos
(acidófilas), o con colorantes básicos (basófilas). Las proporciones son las siguientes:
Células cromófobas (sin gránulos) 50%
Células acidófilas (con gránulos acidófilos) 40%
Células basófilas (con gránulos basófilos) 10%
La parte intermedia está compuesta por células sin gránulos.
Al lóbulo posterior lo forman células nerviosas y gran cantidad de fibras nerviosas. Las fibras
se prolongan dentro del tallo hasta alcanzar el hipotálamo, con el que establecen amplia
comunicación.
Completa la estructura de la hipófisis, un rico sistema de vasos sanguíneos.
Por razones didácticas se comenzará el estudio de la hipófisis por el lóbulo posterior,
invirtiendo así el orden clásico de los textos de anatomía y fisiología.
EL LOBULO POSTERIOR
La parte nerviosa de la hipófisis se prolonga, como se describió, a través del tallo de la
hipófisis, hasta la parte vecina del cerebro, el hipotálamo. Lóbulo posterior, tallo y parte
relacionada del hipotálamo, constituyen una unidad anatómica y funcional.
Se le reconoce a este lóbulo una función bien demostrada, que es la de regular la cantidad de
orina que filtra por el riñón. A tal objeto, en este lóbulo se elabora una sustancia que pasa a la
sangre y llega al riñón, frenando su producción de orina. Esta sustancia, llamada hormona
antidiurética, es fácil de extraer si se preparan extractos de lóbulo posterior, pero su
composición química no es bien conocida.
Cuando espontáneamente se lesiona el lóbulo posterior o se lo extirpa por una operación
quirúrgica, deja de producirse la hormona antidiurética y aparece la diabetes insípida. Se
obtiene el mismo resultado cortando el tallo o destruyendo la parte del hipotálamo en la que
se originan las fibras nerviosas que van por el tallo al lóbulo posterior.
Los pacientes con diabetes insípida orinan cantidades extraordinarias que alcanzan a 10, 20,
incluso 40 litros diarios, y están sometidos a la doble tortura de una sed insaciable y una
necesidad frecuente de la micción. La orina es de color claro, y se diferencia de la orina de la
verdadera diabetes porque no contiene glucosa.
Si se da al enfermo lóbulo posterior de hipófisis desaparecen transitoriamente los síntomas de
la diabetes insípida, por lo que hay que repetir continuamente el tratamiento. En el hombre se
pueden inyectar extractos de este lóbulo o, simplemente, hacer inhalaciones nasales de polvo
del mismo, como si fuera rapé.
Se le atribuyen también al lóbulo posterior, además de la acción antidiurética, otras dos
funciones que todavía no son bien conocidas. Una de ellas sería favorecer en la mujer la
producción del parto, tesis en parte confirmada por el hecho de que se preparan extractos de
este lóbulo capaces de inducirlo.
La otra acción consistiría en regular la presión arterial, probada por Orlas, de la Argentina, en
los batracios. Pero en los mamíferos faltan pruebas suficientes.
LA PARS INTERMEDIA
Es la más pequeña de las 3 partes de la hipófisis, y por estar muy adherida al lóbulo posterior
es dificilísimo separarla, y por ello en los extractos de este último suelen aparecer acciones
agregadas debido a impurezas constituidas por la presencia de trozos del lóbulo intermedio.
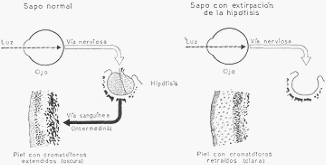
En los mamíferos, la pars intermedia no tiene función conocida; en los batracios regula el color
de la piel. El mecanismo es el siguiente: en el animal normal, la luz impresiona la retina y por
vía nerviosa regula la secreción de una hormona del lóbulo intermedio llamada intermedina,
la que pasa a la sangre y llega a la piel, actuando sobre los cromatóforos, que son células que le
dan color. Si se interrumpe el mecanismo en una de sus partes, por ejemplo extirpando la
hipófisis a un sapo, se suprime la producción de intermedina, y su piel empalidece en pocas
horas volviéndose de un color gris claro; si se le inyecta después intermedina, adquirirá en
pocos minutos un tono carbón.
Representación esquemática del mecanismo que preside los cambios de coloración en la piel
de los batracios. A la izquierda, en un sapo normal; a la derecha, en otro con extirpación de la
hipófisis.
Se llaman cromatóforos unas células que existen en la epidermis de los peces y batracios, que
contienen gran cantidad de gránulos de color negro, rojo, etc. Cuando los gránulos están
dispersos en toda la célula forman una cortina que obstruye el paso de la luz y el animal
aparece del color del gránulo; cuando los gránulos se concentran en un punto de la célula, los
rayos luminosos pueden pasar y la piel del animal adquiere un tono claro.
La intermedina, al llegar a la piel, dispersa los gránulos y la piel del animal se oscurece. Por el
contrario, cuando falta, la piel se aclara.
EL LOBULO ANTERIOR
Este lóbulo tiene múltiples e importantísimas funciones que hacen de él, y con razón, el centro
de la constelación endocrina del organismo.
A) HORMONAS. Uno de los problemas más complejos y debatidos en la actualidad, es el de
conocer las hormonas anterohipofisarias.
Su número exacto se desconoce. En los extractos aparecen numerosas acciones fisiológicas,
producidas quizás artificialmente por rotura de algunas hormonas durante las manipulaciones
químicas de preparación. Dicho en otros términos, la glándula produciría menos de las que se
describen, a juzgar por la acción de los extractos.
Las que se dan como mejor demostradas son: la de crecimiento; la prolactina; la adrenotrófica;
la tirotrófica, y
las dos gonadotróficas. Las obtenidas en estado muy impuro y sobre cuya
individualidad existen dudas son: la diabetógena; la cetogenética; la paratirotrófica; la
mamógena, etc.
En resumen, no se conoce todavía el número exacto de las hormonas hipofisarias, ni tampoco
la fórmula química de las mismas. Las dificultades en estos estudios radican principalmente en
el hecho de que todas estas hormonas son proteínas, y es muy difícil trabajar en la química de
estas sustancias.
B) REGULACION. El lóbulo anterior recibe estímulos que regulan su funcionamiento por dos
vías diferentes: nerviosa y sanguínea.
Hay varias pruebas de que el sistema nervioso 'modifica el funcionamiento de esta parte de la
hipófisis. Mencionaremos que en algunas aves de los países muy fríos y en los hurones, se
produce durante el invierno una atrofia marcada de sus testículos, y que al llegar la primavera,
estos órganos aumentan 1.000 o más veces su tamaño. Esto se explica porque la mayor
iluminación que acompaña a la primavera estimula los ojos y origina un reflejo que, por los
nervios ópticos, llega al cerebro, y de aquí a la hipófisis anterior, donde provoca la secreción
de la hormona estimulante que agranda al testículo. Basta interrumpir el camino de estos
estímulos en cualquier parte de su trayecto, por ejemplo ceguera a causa de una lesión ocular,
para que se observe la desaparición de dicho mecanismo. Una explicación análoga la brinda el
hecho, conocido por los granjeros, de que las gallinas ponen más huevos si se ilumina día y
noche el gallinero. En el hombre se sabe que lesiones de ciertas partes del hipotálamo (por
proyectil, etc.) dan adiposidad y atrofia sexual.
Basten los ejemplos enumerados para demostrar que el sistema nervioso modifica la actividad
del lóbulo anterior. Lo curioso es que, hasta el presente, no se han podido observar al
microscopio los nervios que desde el cerebro penetran en este lóbulo.
Por vía sanguínea llegan a la antero hipófisis hormonas fabricadas en distintas glándulas
endocrinas, que moderan su funcionamiento. Las bien conocidas nacen en las glándulas
sexuales, las suprarrenales y la tiroides. Al tratar cada una de estas glándulas se insistirá en
particular.
Enanismo por extirpación de la hipófisis. Arriba, cachorros hermanos a los 8 días de operar a
dos de ellos; abajo, a los 124 días de la operación. El mayor, sin operar, sirve de testigo para la
prueba
C) FUNCIONES. Para su mejor comprensión se pueden clasificar, siguiendo a Houssay, en los
siguientes grupos: a) de crecimiento; b) metabólicas; c) de regulación de otras endocrinas, y,
separadas de éstas por su importancia, d) las sexuales.
a) Crecimiento. La antero hipófisis regula el crecimiento por intermedio de una hormona que
vierte en la sangre y que Evans, en los Estados Unidos, acaba de obtener en estado de pureza,
bajo la forma de polvo.
Si a un animal en crecimiento se le extirpa la hipófisis, deja de crecer. Se obtienen así animales
enanos que tienden a engordar, si son perros, o a enflaquecer notablemente, si son ratas, y que
resultan muy sensibles al ayuno o a las infecciones. Basta inyectarles la hormona para que
reinicien su crecimiento, o lo sobrepasen, pudiéndose obtener por este camino verdaderos
gigantes de esa especie. Igual resultado se observa en animales normales.
En el hombre es dado observar enanos por causa hipofisaria, semejantes a niños
por su
aspecto, y gigantes como se acostumbra a ver en los circos, cuando por tumores funciona en
exceso.
Si la hiperfunción del lóbulo anterior se produce en un hombre adulto, el crecimiento óseo ya
no es posible sino en la cabeza, las manos y los pies, que adquieren proporciones exageradas;
pero la talla no se altera, constituyendo la enfermedad llamada acromegalia.
b) Metabólicas. Interviene la antero hipófisis en la regulación del consumo de sustancias
nutritivas y en el desprendimiento de calor por las mismas durante sus procesos de
degradación. Pero el mecanismo íntimo por el cual lo hace, se conoce muy imperfectamente y
no se han aislado puras las hormonas que intervienen.
El consumo de las grasas, las proteínas y los hidratos de carbono, es regulado por esta
glándula. Los brillantes estudios de Houssay, en la Argentina, han permitido conocer muchos
de estos puntos, y particularmente el importante papel que la antero hipófisis desempeña con
respecto a los glúcidos, sobre lo que volveremos a propósito del páncreas.
Este investigador ha preparado un extracto de lóbulo anterior, llamado diabetógeno, que
inyectado a perros les provoca una diabetes típica. Esta diabetes hipofisaria es análoga a la que
presentan en el 33% de los casos los enfermos de acromegalia, los cuales padecen o han
padecido de hiperfunción hipofisaria. Houssay, Smyth y Foglia han podido comprobar que las
hipófisis
de cadáveres humanos tienen más sustancia diabetógena que las de otras especies
animales.
Por el contrario, la falta del lóbulo anterior disminuye la producción y el consumo de glúcidos;
así se explica el extraordinario hecho, observado por Houssay en 1930, de que la extirpación
del lóbulo anterior mejora la diabetes, enfermedad en la que se hallan aumentados la
producción y el consumo de estas sustancias.
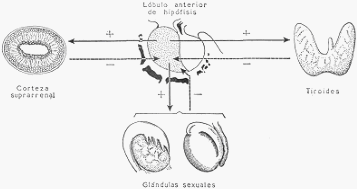
c) Regulación endocrina. El lóbulo anterior de la hipófisis estimula continuamente la corteza
suprarrenal, la tiroides y las gonas (testículo y ovario), y permite que todas estas glándulas
tengan estructura y función normales. Esto lo realiza por hormonas específicas para cada
glándula, que vuelca en la sangre. Estas son: la córticotrofina, la tirotrofina y las
gonadotrofinas. Ya se ha dicho que éstas, por secreciones propias, moderan la hipófisis.
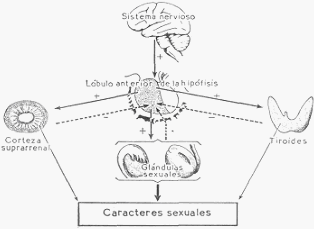
Esquema de las relaciones hormonales entre el lóbulo anterior de la hipófisis y otras glándulas
endocrinas. El signo + indica estimulación, y el signo — inhibición.
En los casos de tumores que destruyen este lóbulo hipofisario, o de extirpaciones quirúrgicas,
faltan las hormonas, y las glándulas correspondientes disminuyen de tamaño rápidamente.
Como dato ilustrativo se puede señalar, que en la rata hipofisopriva hemos observado, a los 15
días de la operación, reducción de los testículos al 10% de su peso inicial, y en las demás
glándulas atrofias tan marcadas, que son perceptibles a simple vista. Además de estos
trastornos anatómicos, se puede demostrar que funcionan menos que antes.
La insuficiencia del lóbulo anterior de hipófisis es restaurada por inyección de extractos del
mismo origen, que corrigen todas las deficiencias. Se obtiene igual resultado por inyección de
una u otra de sus hormonas, pero en este caso el restablecimiento queda limitado a las
propiedades de la sustancia usada. Por inyección de una cantidad excesiva de una hormona,
se puede provocar enorme tamaño e hiperfunción de la glándula por ella estimulada.
Páncreas
El conocimiento del páncreas como glándula endocrina se halla jalonado por dos hechos de
importancia capital en Medicina: el primero fue señalado en Alemania por Von Mering y
Minkowski, en 1889, y es que la extirpación quirúrgica del páncreas, en el perro, conduce
fatalmente a la diabetes, lo que fue la primera demostración de una enfermedad humana
reproducida artificialmente en los animales; el segundo hecho, el sensacional descubrimiento
de la insulina en extractos de páncreas, debido a los investigadores canadienses Banting y
Best, en 1922, significó un incalculable beneficio en el tratamiento de la diabetes humana.
El papel fundamental del páncreas endocrino consiste en mantener dentro de cifras normales
la glucosa del organismo y regular todas sus transformaciones, sean éstas su absorción por vía
digestiva, su producción, su depósito o su consumo. Cumple estas variadas funciones por
intermedio de la insulina, activa hormona que vierte en la sangre.
ANATOMIA
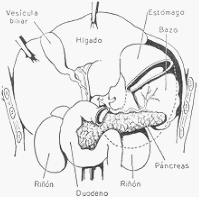
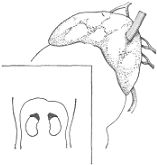
El páncreas del hombre es una glándula abdominal de forma comparable a la de un martillo,
cuya cabeza se adhiere firmemente al intestino en su salida del estómago (duodeno), y su
cuerpo se extiende horizontalmente hacia la izquierda hasta tocar el bazo. Mide unos 16 a 20
cm de largo, 4 a 5 cm de alto y 2 a 3 cm de espesor en su cuerpo, y pesa alrededor de 70
gramos. Su color es rosado.
Páncreas en el abdomen, visto por delante, luego de levantar el hígado y extirpar el estómago.
Su estructura es compacta y se halla recorrido por conductos que confluyen entre sí y terminan
formando dos que son principales y que desembocan en el duodeno. Por estos conductos corre
el jugo pancreático que representa la secreción externa del órgano, la que en la luz del intestino
digiere las grasas, las proteínas y los glúcidos.
Examinándolo al microscopio se observa innumerable cantidad de vesículas, con una capa de
células en su pared, y los conductos ya señalados, que nacen en las vesículas. Las células de
éstas son las productoras de la secreción externa, que sigue luego por los conductos hacia el
duodeno. Desparramadas, aquí y allá entre las vesículas, se hallan unas pequeñas y escasas
formaciones, descritas por primera vez por Langerhans en 1862, y llamadas hoy, en homenaje
a su descubridor, islotes de Langerhans.
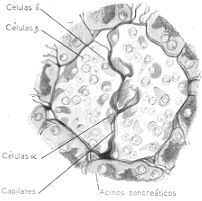
Los islotes de Langerhans son irregularmente. Están formados por células de 5 variedades
diferentes, apoyadas las unas en las otras y rodeado el conjunto por una delgada cápsula. De
estas células las designadas células beta (p) son las productoras de la secreción interna del
páncreas, hormona bautizada con el nombre de insulina por provenir de las ínsulas o islotes
pancreáticos.
En resumen, el páncreas es una glándula mixta con una secreción externa: el jugo pancreático;
y una secreción interna: la insulina. Aquí sólo se estudiará la secreción interna.
LA INSULINA
La insulina es la hormona de secreción interna del páncreas. La demostración de este hecho y
el aislamiento químico de la insulina, bien confirmados hoy, han sido objeto de vivas
polémicas y de enormes esfuerzos.
A) ORIGEN.
La insulina es producida por las células beta de los islotes de Langerhans del
páncreas.
Islote de Langerhans y células que lo componen. Las B elaboran insulina.
Estos hechos lo demuestran: 1) se la obtiene de extractos de páncreas en los que se ha
destruido el páncreas exocrino, productor del jugo pancreático; 2) en el hombre se observan
tumores formados exclusivamente por células beta, de los que se puede extraer insulina; 3) en
los casos de diabetes humana hay muchas células beta destruidas; 4) la destrucción de las
células beta, por inyección de aloxana, o de extractos de lóbulo anterior de hipófisis (Houssay),
produce diabetes.
B) QUIMICA. La insulina es una sustancia de naturaleza proteica, cuya composición química
exacta se ignora todavía, aunque se sabe que contiene carbono, hidrógeno, oxígeno, nitrógeno
y azufre.
Comercialmente se la prepara extrayéndola del páncreas de vaca, de cerdo o de oveja, y hasta
la fecha no ha sido producida artificialmente en el laboratorio. Su grado de pureza llega a ser
tal, que se la puede obtener como cristales visibles al microscopio.
En medicina se la utiliza bajo dos formas: la) insulina regular, de efecto rápido y pasajero, y 23)
insulina asociada a protamina, u otras sustancias similares, de efecto lento y prolongado. Cada
una de estas variedades tiene su particular aplicación terapéutica.
C) ACCION. En el sujeto normal el efecto más notable es la disminución de la glucosa de la
sangre, que baja del nivel de 1 g por litro hasta cifras vecinas a 0,30 g por litro, según la dosis
inyectada. Al acercarse a esta cifra, se producen varios síntomas, como sudor, molestias
gástricas, mareos, etc., y al final violentas convulsiones de todos los músculos, que preceden a
la muerte si no se inyecta glucosa. Esta acción hipoglucemiante se aprovecha para medir la
insulina, y se llama unidad la dosis necesaria para bajar a ese nivel la glucemia de un conejo, y
que consiste en de la dosis anterior.
La disminución de la glucosa sanguínea se explica porque una parte se deposita en los
músculos formando glucógeno, del que aumenta la cantidad, y el resto es quemado y
desaparece.
En el diabético tiene las mismas acciones que en el hombre normal y se deposita la glucosa en
forma de glucógeno en el hígado, donde está disminuida. Además, corrige las alteraciones
debidas a esta enfermedad.
REGULACION DE LA SECRECION DE INSULINA
El estímulo específico que hace segregar insulina es el tenor de glucosa en la sangre.
La glucosa estimula directamente los islotes de Langerhans, como puede probarse
enriqueciendo en glucosa la sangre que llega al páncreas, y observando la descarga de
insulina, que se produce aunque se destruyan totalmente los nervios pancreáticos (Foglia). A
la acción directa de la glucosa sobre la secreción de insulina, se suma una leve acción
estimulante indirecta que obra sobre los centros nerviosos que por los nervios envían
estímulos a los islotes. Por los dos mecanismos, sanguíneo y nervioso, la glucosa asegura al
organismo una provisión de insulina permanente y adecuada a sus necesidades.
La secreción de insulina por el páncreas varía de un momento a otro, según se deduce de lo
señalado. El mínimum se obtiene cuando el organismo está en reposo y en ayunas, y es para
un adulto normal de unas 50 unidades por día. Naturalmente, aumenta con el ejercicio y,
sobre todo, después de una alimentación rica en azúcar.
El páncreas posee gran adaptabilidad
para segregar insulina. Lo prueba el hecho de que un animal permanece normal, tanto
reduciendo su masa pancreática al%o, por extirpación quirúrgica, como aumentándola 4 veces
por injerto; es decir, que tolera variaciones de su masa productora de insulina como 1:40, y las
compensa aumentando su secreción de insulina si se reduce el páncreas, o a la inversa
disminuyendo la secreción si aumenta, de modo de segregar siempre la cantidad necesaria.
FUNCIONES
El páncreas, por medio de la secreción de insulina que vierte en la sangre, mantiene normal el
azúcar sanguíneo o glucemia, y regula la producción y consumo del mismo
Tanto la disminución de la secreción de insulina como su exceso causan graves trastornos, bien
conocidos en medicina, ya que hay enfermedades que azotan a la humanidad que los
provocan en forma espontánea.
A) HIPOFUNCION: DIABETES. La diabetes se produce espontáneamente en el hombre. En
los animales de laboratorio se la obtiene por extirpación quirúrgica del páncreas o su lesión
por medios químicos como la aloxana, o por inyección de extracto diabetógeno de lóbulo
anterior de hipófisis (Houssay). En todos los casos los islotes se hallan lesionados o ausentes, y
la secreción de insulina disminuida o desaparecida.
Los síntomas observados aparecen más o menos bruscamente, son más o menos intensos
según el caso, pero siempre se nota: gran sed, gran apetito (los enfermos son grandes
comilones); a pesar de eso se nota enflaquecimiento y debilidad progresivas, facilidad para las
infecciones, aumento de la cantidad de orina y, finalmente, la muerte, precedida a veces por
un estado de inconsciencia, el coma diabético. Los análisis revelan un exagerado aumento de
azúcar en la sangre (2, 3, 4 o más gramos por mil) y su aparición en la orina, disminución de
sus reservas en el hígado, y dificultad en su consumo. Se agregan alteraciones profundas en el
contenido de grasas, proteínas y sus derivados en la sangre y orina. Es decir, que en la diabetes
hay un profundo trastorno en la nutrición de todas las sustancias y particularmente de los
azúcares.
El cuadro anterior se corrige inyectando insulina, regulando la dieta o, como lo demostrara
con espectaculares experimentos el fisiólogo argentino Houssay, extirpando el lóbulo anterior
de la hipófisis (animales de Houssay).
B) HIPERFUNCION: HIPERINSULINISMO. Se puede provocar inyectando insulina en dosis
grande, o también se lo observa espontáneamente como enfermedad.
La inyección de insulina trae la hipoglucemia, que ya se explicó, y los síntomas graves
acompañados de convulsiones, que si no se corrigen a tiempo, dando glucosa, llevan a la
muerte. Este procedimiento se aplica para curar algunas enfermedades mentales, por la fuerte
conmoción que provoca. Dando cantidades pequeñas de insulina, aumenta el apetito y el
sujeto engorda.
El hiperinsulinismo, o exceso de insulina producido espontáneamente, se observa en el curso
de tumores de las células beta del páncreas. Los sujetos atacados de esta enfermedad
presentan, por el ayuno, crisis de hipoglucemia con sudores, hambre, mareos y, en los casos
graves, convulsiones e incluso llegan a la muerte; sólo pueden ser aliviados por ingestión de
azúcar y curados por la extirpación del tumor.
Suprarrenales
Las glándulas suprarrenales son indispensables para la vida. Se las puede considerar como las
glándulas de la sal, ya que gobiernan los intercambios de ésta dentro del organismo.
ANATOMIA
Son dos, ubicada una sobre el polo superior del riñón derecho y otra sobre el izquierdo. Tienen
la forma y el tamaño de un gajo de naranja pequeña. Miden 30 mm de alto, 25 mm de ancho y
7 mm de espesor. Cada una pesa 5,6 gramos.
Relación de una glándula suprarrenal con el polo superior del riñón. En pequeño, y en negro,
situación de las dos en el abdomen.
Al microscopio se observa que cada una de ellas tiene dos partes, de distinta estructura, que
contienen dos glándulas endocrinas diferentes: la corteza, compuesta por células que nacen en
la parte más externa y, a medida que envejecen, se cargan de gránulos de grasa y se hacen más
internas; y la medula, situada con respecto a la otra como el carozo a la pulpa del melocotón,
formada por otro tipo de células, que se tiñen electivamente con el ácido crómico, y que por
eso se las llama cromafinas.
Gran número de nervios llegan, sobre todo a la medula, y también muchos vasos sanguíneos,
al punto que es la más irrigada de todas las glándulas endocrinas.
MEDULA SUPRARRENAL
Se puede vivir casi perfectamente sin las dos medulas suprarrenales; Houssay y Lewis lo han
demostrado, por medio de singulares experiencias, en forma concluyente.
La medula elabora una sustancia llamada adrenalina, que tiene el mérito
histórico de ser la
primera hormona cuya fórmula química se descubrió (por Takamine y simultáneamente
Aldrich). Hoy día se la prepara por extracción de las suprarrenales, o bien en forma sintética
en el laboratorio, y sus preparados se hallan en el comercio.
La secreción de adrenalina se halla bajo el control del sistema nervioso: si se excitan con una
corriente eléctrica los nervios que van a la glándula, se produce una descarga de adrenalina a
la sangre; si se los corta se reduce la secreción a niveles mínimos. Este mecanismo nervioso
entra en acción en diversas circunstancias de la vida diaria, y produce la descarga de
adrenalina cuando el organismo pasa por una situación de apremio, como, por ejemplo,
durante el miedo o furor intenso, frío excesivo, ejercicio muscular, pérdida grande y brusca de
sangre, etc.
La inyección de adrenalina produce los mismos efectos que la excitación del sistema nervioso
simpático (acción simpático mimética). Las acciones principales de la adrenalina son:
contracción de las arteriolas, que provoca una fuerte pero transitoria elevación de la presión
arterial; aumento de la glucosa en la sangre y escape consecutivo por la orina; relajación de los
músculos bronquiales.
Por estas acciones se la utiliza en terapéutica. Por su acción constrictiva, para cerrar los
pequeños vasos y detener las hemorragias en la nariz, la boca, etc.; por su acción hipertensora,
para elevar la presión arterial en casos de síncope, donde ha descendido bruscamente; por su
acción hiperglucemiante, para aumentar el azúcar de la sangre en los casos en que ha
disminuido en forma peligrosa, como luego de inyecciones grandes de insulina. También se la
utiliza para aliviar en caso de ataque de asma, y permitir una respiración fácil, porque relaja
los músculos que cierran los bronquios finos y dificultan el pasaje de aire para dentro o fuera
del pulmón.
CORTEZA SUPRARRENAL
A la inversa de la medula, la corteza es indispensable para la vida; por eso se dice que tiene
papel vital. Los animales privados de ella mueren en un plazo de días, que es variable según la
especie:
Perros; sapos 2-4 días
Gatos; cobayos: 6-10 días
Ratas: hasta 30 días.
Un octavo de la suma de las dos cortezas basta para proteger de la muerte. Algunos animales
sobreviven a la extirpación de las suprarrenales debido al hecho que poseen suprarrenales
accesorias en otras partes. Esto se observa en algunas ratas.
A) HORMONAS. Todavía no se conocen con seguridad las hormonas elaboradas por la
corteza. Los químicos han aislado unas 28 sustancias, pero se sospecha que algunas son
productos elaborados en el curso de la extracción.
Todas estas sustancias se llaman esteroides, tienen una fórmula química bastante parecida y
derivan de un núcleo químico común. Se hallan emparentadas con las hormonas producidas
por las glándulas sexuales, con algunas sustancias que se encuentran en la bilis (colesterol), ya
que todas tienen composición química parecida.
De los esteroides suprarrenales se destaca la desoxicórticosterona —muy utilizada en
Medicina en forma de acetato de desoxicórticosterona, llamada abreviadamente DOCA—, la
que por inyección es muy activa y hace que el organismo retenga la sal. Otros de los esteroides
son iguales a los elaborados por el testículo y el ovario, y son capaces de masculinizar o
feminizar al sujeto en que se inyectan.
Los extractos brutos de corteza suprarrenal contienen una mezcla de los esteroides señalados y
otras sustancias desconocidas por ahora. Son sumamente activos.
Los esteroides de la corteza terminan eliminándose por la orina, donde reciben el nombre
general de 17cetoesteroides. Aquí se los mide y se juzga, por su cantidad, cómo funciona la
glándula. Se hallan en exceso en las hiperfunciones, corno, por ejemplo, cuando existe algún
tumor suprarrenal, y disminuidos en casos de destrucción de la corteza.
B) REGULACION. El lóbulo anterior de hipófisis gobierna a la corteza suprarrenal por medio
de una hormona que envía por la sangre, la córticotrofina. El exceso de córticotrofina agranda
las suprarrenales y su falta las reduce y disminuye su función.
A la izquierda, corte de una suprarrenal de rata normal; a la derecha, de una rata sin hipófisis.
Ambos cortes son vistos con el mismo aumento.
La corteza, a su vez, por intermedio de sus hormonas que vuelca en la sangre, modera la
función hipofisaria.
De ello resulta que la hipófisis y la suprarrenal se regulan recíprocamente gracias a un juego
hormonal que tiene lugar en la sangre.
C) FUNCIONES.
Por medio de sus hormonas regula la distribución de la sal y el agua, el
consumo de los azúcares, y vuelve al organismo resistente contra diversos agentes nocivos. En
ciertas condiciones, pone de manifiesto su influencia en la esfera sexual.
Este conjunto de funciones, tan diversas, se aprecia muy bien en los casos en que la glándula
es lesionada por enfermedad o experimentalmente. La enfermedad de Addison, descrita
magistralmente por este médico inglés, ilustra en forma clara sobre los síntomas de
insuficiencia de la corteza suprarrenal en el hombre, que son análogos a los observados en los
animales de laboratorio.
a) Hipofunción: Enfermedad de Addison. Esta enfermedad se inicia por tuberculosis, cáncer,
etc., que ataca y destruye la glándula. Luego el enfermo enflaquece progresivamente, su piel
cobra un tinte bronceado que solamente lo muestra la especie humana, tiene malestares
digestivos y una extraordinaria pérdida de fuerza (astenia), que llega a extremos tales, que lo
imposibilitan para llevar una cuchara a la boca. Finalmente, si el individuo no es tratado,
muere después de un año o más de padecimientos.
Si se examina con medios adecuados a estos pacientes, se comprueba su dificultad para retener
dentro de la sangre la sal y el agua, que escapan por la orina y, naturalmente, disminuyen en
la sangre. Por este motivo la sangre se concentra y disminuye en volumen, la presión arterial
cae y el sujeto pierde peso. Basta agregar a la dieta sal en exceso, para que desaparezcan todos
estos síntomas y el sujeto pueda sobrevivir.
Si se continúa el examen se observa que el hígado y la sangre tienen poca glucosa y la pierden
fácilmente a causa del ayuno o del ejercicio, llegando hasta la muerte si desciende mucho y no
es restaurada a tiempo ingiriendo o inyectando azúcar. Por esta acción depresora, la
extirpación suprarrenal resulta beneficiosa en la diabetes experimental, porque hace descender
el azúcar aumentado, aunque aparecen otros síntomas de insuficiencia suprarrenal.
Finalmente se comprueba lo débil que son las resistencias orgánicas de estos individuos contra
los más diversos agentes. Así una sangría, la exposición al frío, una infección, que serían bien
toleradas por un sujeto sano, provocan en estos pacientes graves trastornos o la muerte misma.
Todos los síntomas anteriores producidos por insuficiencia de las hormonas suprarrenales,
comprobados además por
la disminución de cetoesteroides en la orina, se corrigen con la
inyección de extractos totales de esta glándula o algunas de sus hormonas. Como los
trastornos internos debidos a la pérdida de agua y sal son los más graves del conjunto, basta
tratarlos para obtener un resultado feliz. Esto se obtiene aumentando la ingestión de sal en la
comida, o inyectando la hormona Doca que hace retener al organismo la sal y el agua.
b) Hiperfunción. Es conocida casi exclusivamente en la especie humana. En el caso de ciertos
tumores de las suprarrenales se observa una producción exagerada de hormonas masculinas,
que, cuando aparecen en mujeres adultas las masculinizan, en el sentido que aumenta su
fuerza muscular, les aparecen pelos en el cuerpo junto con barba y bigote, y también diabetes.
Casos como estos se ven a veces en los circos.
Si tumores de este tipo se producen en los niños, aparece un desarrollo general y sexual precoz
y exagerado, dando los llamados "niños Hércules".
La extirpación de los tumores hace desaparecer los cuadros señalados.
Tiroides
Distinguen a esta glándula dos características fundamentales: 1) su íntima vinculación al yodo,
y 2) su propiedad de activar las combustiones orgánicas.
ANATOMIA
Está formada por dos lóbulos verticales, unidos por un istmo, que dibujan en conjunto la letra
H; el conjunto está situado por delante de la tráquea, en la base del cuello. Su altura es de unos
3 cm, su ancho de 6 a 7 cm, y su espesor de 20 a 5 mm. Su peso es de 20 a 35 gramos, pero es
corriente observar tamaños muy variables. En la mujer es mayor que en el hombre.
Posición de la tiroides en el cuello, sobre la tráquea, después de abrir la piel y los músculos
que la cubren.
Una rica inervación e irrigación sanguínea son factores que hablan de su importancia
fisiológica.
Examinada al microscopio se la ve constituida por numerosas celdillas redondeadas,
completamente cerradas, que se tocan las unas a las otras como las uvas de un racimo. La cara
interna de estas celdillas se halla revestida por una capa de células cúbicas, y el resto de la
cavidad se halla rellenado por una sustancia gelatinosa (coloide).
La celdilla se llama folículo tiroideo, mide unos 300 p. de diámetro y constituye la unidad
anatómica y funcional de esta glándula. La sustancia gelatinosa es la secreción tiroidea,
contiene la hormona y es producida por las células de la pared.
Tiroides de sapos: a la izquierda, sin hipófisis; al centro, normal; a la derecha, inyectado con
lóbulo anterior de hipófisis.
EL YODO Y LA TIROIDES
El yodo es un metaloide que existe en el suelo. De allí lo arrastra el agua y lo absorben las
plantas. Los animales, al alimentarse lo toman de estas fuentes y lo conservan en su organismo
El hombre ingiere yodo con el agua
que bebe y las plantas y animales que come, en una
cantidad de 100 a 200 microgramos diarios. El microgramo es igual a una millonésima de
gramo.
Después de absorbido, el yodo circula por la sangre en estado de yoduro y de allí lo toma la
tiroides con una avidez extraordinaria, superior a la de cualquier otro órgano, y lo concentra,
dentro de ella, aproximadamente 10.000 veces más que en la sangre. La tiroides contiene así
del yodo total del organismo.
Las células foliculares fabrican la hormona tiroidea uniendo por un complicado proceso el
yodo ingerido a una proteína. Parte de esta hormona se deposita en el coloide intrafolicular
bajo la forma de un compuesto de molécula gigante, llamado tiroglobulina, aislado por
Ostwald en 1899. El resto pasa a la sangre y se reparte por todo el organismo, también bajo la
forma de un compuesto yodado, que todavía no ha sido bien identificado.
Hay sustancias capaces de modificar el proceso de formación de la hormona tiroidea y su
conocimiento interesa al médico. El tiouracilo y otras sustancias emparentadas con éste lo
traban, disminuyendo la formación de la hormona, lo que es útil para tratar a los que la
fabrican con exceso (hipertiroideos). El exceso de yodo en la dieta también lo modera y es
aprovechado con igual objeto; la falta de yodo trae alteraciones graves que trataremos en
seguida.
Finalmente el yodo ya utilizado encuentra su vía de salida por el intestino, el sudor y, sobre
todo, por la orina.
REGULACION. BOCIO
Hay dos influencias fisiológicas que regulan el funcionamiento de la tiroides, son la hipófisis y
el yodo.
El lóbulo anterior de la hipófisis estimula a esta glándula por la tirotrofina. Esta sustancia pasa
continuamente a la sangre, llega a la tiroides, y desempeña el papel del látigo con respecto al
caballo. Cuando falta se altera la imagen histológica de la tiroides y ésta funciona menos, y a la
inversa, por un exceso se puede obtener figuras y signos de hiperfunción.
Por otra parte, la secreción tiroidea pasa a la sangre, llega a la hipófisis y la mantiene frenada,
lo que se comprueba porque al extirpar la tiroides, la hipófisis funciona con exceso.
El yodo ingerido, que llega por la sangre, es el otro gran regulador de la actividad tiroidea. Un
exceso del mismo modera a la glándula, pero un déficit también trae graves trastornos,
conocidos desde la más remota antigüedad con el nombre de bocio.
Existen grandes zonas en nuestro planeta que contienen poco yodo en sus tierras.
Mapa de la repartición mundial del bocio.
Los hombres y animales de esas zonas ingieren diariamente cantidades de yodo inferiores a
100 ó 200 microgramos, y en ellos, con el andar del tiempo, la glándula tiroides empieza a
aumentar de tamaño, hasta adquirir a veces proporciones monstruosas y deformar la base del
cuello; a esta hipertrofia tiroidea se la denomina bocio.
En las zonas bocígenas no sólo más del 10 al 20% de sus habitantes tienen bocio perceptible a
simple vista, sino que aumenta en forma pavorosa la cantidad de tarados de todo orden y
sobre todo mentales: sordomudos, paralíticos, retardados, cretinos y otros trastornos
psíquicos, y hay numerosos índices que demuestran en los restantes sujetos, aparentemente
sanos, una disminución del vigor de la raza (menor talla, menor diámetro torácico, menor peso
corporal, etc.). Por esta razón es más exacto llamar endemia bociocretínica, a la existencia de
bocio en una zona.
Mujer con bocio gigante que deforma el cuello.
Para hacer desaparecer la endemia bociosa hay que dar a ingerir a los sujetos de la zona el
yodo que no reciben con sus alimentos. Países como Suiza, Inglaterra, Estados Unidos, etc.,
han hecho disminuir o desaparecer el bocio por el agregado de yodo a la sal de cocina, en una
proporción aproximada de 100 mg de yodo por kilo de sal.
En la Argentina existe bocio en toda la zona cordillerana y en provincias alejadas de la misma,
como la de Córdoba o la gobernación de Misiones. Los esfuerzos para evitarlo son
insuficientes y todavía no se han aprovechado los beneficios incalculables de la sal yodada.
FUNCIONES
La tiroides, por intermedio de su secreción, estimula la nutrición de las células, que se revela
particularmente por su influencia marcada en el crecimiento del individuo, el
desprendimiento de calor y el mantenimiento normal de la actividad psíquica. Además, en los
batracios regula la metamorfosis.
Estos hechos pueden ser bien demostrados provocando en un animal de experimentación la
función insuficiente de la tiroides por su extirpación, o la función excesiva por inyección de
extractos activos.
En el hombre se observan cuadros clínicos semejantes, producidos por la enfermedad, y a ellos
nos referiremos como ejemplo en las descripciones que siguen.
A) INSUFICIENCIA TIROIDEA. Cuando en los primeros años de la vida la glándula tiroidea
es destruida y deja de funcionar, el crecimiento corporal se interrumpe y el sujeto queda
enano.
Enanismo por extirpación de la tiroides en la cabra. A la izquierda, sin tiroides; a la derecha,
normal.
A este trastorno se le suman otros muy especiales y fáciles de reconocer. Sobresale el
mixedema o infiltración de líquido en la piel, que hace a ésta áspera, fría e hinchada, por cuya
razón los dedos son gordos y la cara redondeada, con hinchazón de los párpados. Los pelos
son lacios y suelen caer, siendo escasos, particularmente en las cejas. Acompañan a los
anteriores graves signos de déficit intelectual que pueden hacer del sujeto un retardado mental
o un cretino, incapaz de aprender las cosas más elementales (opas del Norte argentino).
Una medición de la cantidad de calor producido por el enfermo confirma el diagnóstico, al
señalar cifras 30 a 45% inferiores a las normales.
El cuadro anterior se observa también en los adultos, aunque no hay en ellos trastornos del
crecimiento y los trastornos mentales son más atenuados.
Los síntomas de insuficiencia tiroidea se curan o mejoran dando de comer al paciente tiroides
o bien inyectándole extractos de esta glándula, en forma continuada, porque su efecto es
transitorio.
B) HIPERFUNCION TIROIDEA. Los pacientes con síntomas de esta naturaleza, si son jóvenes
pueden tener un crecimiento más rápido que los normales, pero nunca llegan al gigantismo.
Son flacos porque pierden agua fácilmente y además queman sus alimentos en forma
exagerada, lo que se comprueba midiendo la cantidad de calor desprendido que sobrepasa en
50%, 100% y más a las normales.
Sus ojos son grandes, abiertos, salientes, y muestran el blanco, dando a la mirada un aspecto
especial (mirada trágica) que permite reconocerlos al pasar por la calle. Este signo sólo se
observa en la especie humana.
Completan el cuadro síntomas nerviosos variados como el temblor de las manos y del cuerpo,
el insomnio, la excitación psíquica que los hace enojadizos y hasta pendencieros.
Para moderar la tiroides y disminuir su secreción se puede dar yodo en cantidad, o tiouracilo,
o bien efectuar la extirpación quirúrgica de la glándula.
C) METAMORFOSIS EN LOS BATRACIOS. La tiroides también estimula el pasaje del estado
de larva al de individuo adulto en estos animales. Por su extirpación se obtienen larvas
pequeñitas que persisten en ese estado sin metamorfosearse. Por el contrario, agregando polvo
de tiroides al agua donde nadan larvas de rana, éstas se transforman aceleradamente en
minúsculas ranas adultas (Guddernatsch).
Paratiroides
Las paratiroides son glándulas indispensables para la vida. Su verdadera importancia pasó
desapercibida durante mucho tiempo por su pequeño tamaño y su vecindad inmediata a la
tiroides, que ha hecho atribuir a ésta, equivocadamente, síntomas que pertenecían a las
paratiroides.
En efecto, durante la época en que los cirujanos se atrevieron a extirpar bocios gigantescos a
personas, observaron a continuación una grave sintomatología, que atribuyeron a la
extirpación de la tiroides bociosa, caracterizada por graves trastornos nerviosos y musculares a
los que denominaron tetania. Luego se vio que se evitaba la tetania respetando a las
paratiroides y, así, fueron separadas en sus funciones las dos glándulas, tiroides y
paratiroides.
Hoy este último hecho se halla bien confirmado. Se sabe que la paratiroides es la glándula que
regula las transformaciones del calcio y el fósforo en el organismo, y que la perturbación de
esa regulación trae aparejados graves trastornos que se revelan por síntomas nerviosos, óseos,
sanguíneos, etc., que en última instancia conducen a la muerte.
ANATOMIA
En el hombre hay cuatro pequeñas glándulas paratiroides ubicadas en el cuello. De éstas, dos
están sobre el lado derecho, en íntima conexión con el lóbulo derecho de la tiroides, y dos en el
lado izquierdo, en igual relación con el lóbulo izquierdo de la tiroides.
La forma de cada una de las paratiroides es ovalada, y miden aproximadamente 7 x 4 x 2 mm.
El peso de las cuatro llega a 140 miligramos.
Estudiándolas por medio del microscopio muestran una estructura compacta, formada por
dos tipos diferentes de células, que las distinguen netamente del tejido vesiculoso tiroideo
vecino.
HORMONA
Los esfuerzos para extraer de esta glándula el principio activo fueron coronados por el éxito en
los trabajos de Collip, en 1925. Este hombre de ciencia canadiense preparó, partiendo de
paratiroides, un producto activo aunque no completamente puro, que tiene las acciones que se
describen para la glándula. Lo llamó parathormona.
Inyectada a los animales moviliza el calcio de los huesos a la sangre, donde, al aumentar en
forma excesiva, pasa a los tejidos blandos y allí se deposita. La desmineralización de los
huesos, provocada por este mecanismo, facilita sus fracturas y dificulta la curación de éstas; el
depósito en los tejidos blandos trae trastornos que dependen del tejido en que se haga
(corazón, músculos, pulmones, riñones, etc.).
Cuando la parathormona se inyecta a personas con síntomas de insuficiencia de paratiroides
(tetania), contribuye a su curación. •
REGULACION DE LA SECRECION PARATIROIDEA
No se ha demostrado en forma absoluta que la hipófisis gobierne el funcionamiento de las
paratiroides como lo hace con la tiroides, las suprarrenales o las glándulas sexuales. Hay
algunos argumentos en favor de la tesis de que el lóbulo anterior hipofisario vierte en la
sangre una sustancia que al llegar a las paratiroides las estimula, pero todavía no hay pruebas
concluyentes.
En cambio., Houssay y sus colaboradores han demostrado que existe una relación estrecha
entre el contenido de calcio de la sangre y la producción de hormona para tiroidea: ésta
aumenta cuando el caldo baja, y, viceversa, disminuye si el calcio sube. Es decir que el tenor
del calcio sanguíneo es el regulador de la secreción de hormona paratiroidea.
FUNCIONES
El papel de esta glándula en el organismo puede ser bien comprendido si se estudian por
separado las consecuencias de su falta y de su exceso de función.
A) INSUFICIENCIA PARATIROIDEA. Se la observa a raíz de la extirpación quirúrgica de la
glándula. Ya se señaló, al principio, que en el hombre fue un accidente frecuente, y ahora lo es
menos, de las operaciones de bocio.
El cuadro clínico que aparece se llama tetania paratiropriva, y comprende dos tipos: aguda y
crónica.
a) Forma aguda. La forma aguda aparece pocos días después de la operación y puede terminar
en la muerte o pasar al estado crónico. Aparte de un malestar general acompañado de
vómitos, aumento de temperatura corporal, etc., se presentan síntomas nerviosos, musculares
y sanguíneos que son característicos.
El sujeto tiene un débil temblor en los miembros y también rigidez que impone a las manos y a
los pies una actitud especial.
Posición de las manos y de los pies en la tetania producida por insuficiencia de las
paratiroides.
Basta apretar la parte interna del brazo y así comprimir los nervios para ver acentuarse la
rigidez de la mano; golpeando el nervio facial en su pasaje por debajo de la piel y por delante
de la oreja, se producen muecas en la cara. En los niños pueden contraerse los músculos
laríngeos y producirse asfixia, por dificultar el pasaje del aire por las vías aéreas. De tanto en
tanto se observan convulsiones.
En la sangre hay dos hechos llamativos: la disminución del calcio por debajo de la cifra normal
de 10 mg por cada 100 cm3 de sangre, y el aumento del fósforo inorgánico arriba de 5 mg por
cada 100 cm3.
b) Forma crónica. Salvándose de la fase anterior que dura días, la insuficiencia paratiroidea
puede ser tolerada por largo tiempo, aunque existe la amenaza de que se intercale un episodio
agudo en cualquier momento.
En esta fase se observa una descalcificación progresiva de los huesos y los dientes, que se
deforman y fracturan con facilidad, mostrando luego una gran lentitud para su reparación. En
los ojos aparecen cataratas y en la piel úlceras con muy poca tendencia a la curación.
c) Tratamiento. El tratamiento de la insuficiencia paratiroidea consiste, por una parte, en evitar
la influencia de factores que agravan la enfermedad, y por otra, en utilizar la acción benéfica
de sustancias que obran elevando el porcentaje de calcio sanguíneo o disminuyendo la
excitabilidad nerviosa.
Los factores agravantes de la enfermedad son varios: el frío o el calor excesivos, la dieta a base
de carne, etc. Ellos empeoran los síntomas de la forma aguda o los hacen aparecer en la
crónica, razón por la cual deben ser evitados.
Entre los factores benéficos figuran como principales los que aportan calcio y así aumentan el
porcentaje sanguíneo, descendido por la enfermedad. Su efecto es notable y merecen citarse:
una dieta rica en leche, la ingestión de sales de calcio, la inyección de vitamina D, que hace
absorber mucho calcio por el intestino, y la de parathormona, que lo moviliza de los huesos,
como ya se explicó. Cabe agregar entre los factores benéficos a los sedantes del sistema
nervioso, que mitigan la exagerada excitabilidad del mismo (bromuros, etc.).
B) HIPERPARATIROIDISMO. Se observan casos de hiperfunción de las paratiroides: en el
hombre espontáneamente por acción de tumores de estas glándulas, y en los animales de
laboratorio provocados, por inyección excesiva de parathormona. Los síntomas son análogos.
En el caso de los tumores se observa la desmineralización de los huesos y dientes, lo que lleva
a fracturas fáciles, a su difícil curación, y al aumento del calcio sanguíneo. Puede obtenerse la
curación si la extirpación del tumor es precoz y radical.
Funciones de reproducción
La importancia y complejidad de las funciones de reproducción justifican ampliamente que se
dedique un capítulo a su estudio. Los últimos años han
permitido agregar, en este terreno,
nuevos conocimientos que han tenido en seguida insospechadas y beneficiosas aplicaciones en
Medicina. Aunque desde cierto punto de vista este capítulo forma parte de las secreciones
internas, que se acaban de estudiar, su separación es razonable por hallarse estrechamente
vinculadas a él funciones no hormonales, como la producción de los gametos, el crecimiento
del feto, etc.
CARACTERES SEXUALES: DESCRIPCION. DIFERENCIA SEXUAL
En los vertebrados hay animales machos y animales hembras, cada uno con caracteres
sexuales propios. La diferencia entre ambos es fácil o no de establecer, según la especie de
animal de que se trate. Así una simple ojeada basta
para distinguir a un león de una leona, un gallo de una gallina, un ciervo macho de una
hembra, gracias a los atributos externos propios de cada especie como son la melena, el
plumaje, los cuernos, el tamaño, etc. En cambio el diagnóstico de sexo no se realiza con igual
facilidad en muchas aves, víboras, sapos, ranas, etc.
Sin embargo, cualquiera sea el caso, se debe tener presente que el único carácter diferencial de
valor absoluto entre ambos sexos, es la presencia de testículo o glándula masculina, o de
ovario o glándula femenina. Estas glándulas constituyen el carácter sexual primario y todas las
restantes diferencias, los caracteres sexuales secundarios.
Diferencias entre las proporciones corporales del hombre y la mujer; las mismas señalan los
caracteres sexuales secundarios.
En la especie humana los individuos normales adultos presentan una marcada y conocida
diferencia sexual. Los caracteres sexuales secundarios se clasifican en: a) morfológicos, los que
corresponden a sus diferencias anatómicas, genitales o no (órganos genitales, estatura,
desarrollo muscular, barba, etc.); b) funcionales, que son los pertenecientes a sus diferencias
fisiológicas (voz, metabolismo, etc.), y c) psíquicos, los de la esfera espiritual. Sin entrar en
detalles reproducimos un cuadro de Houssay, modificado por nosotros, en el que se anotan
ejemplos de algunas de estas diferencias.
EVOLUCION
Aparte de las diferencias del macho con la hembra que acabamos de mencionar, los caracteres
sexuales se distinguen por un ritmo evolutivo particular, con modalidad propia para cada
sexo. Los órganos y funciones sexuales quedan infantiles desde el nacimiento hasta la edad
llamada de la pubertad, en que más o menos bruscamente sufren un marcado desarrollo, hasta
adquirir la madurez sexual (edad adulta), en el que permanecen con variantes; llegada cierta
edad se produce una declinación más o menos brusca y acentuada según el sexo de que se
trate.
Curva esquemática de la evolución de los caracteres sexuales en el hombre y la mujer. El trazo
punteado corresponde a la evolución masculina, el trazo lleno a la femenina
En el hombre la pubertad aparece entre los 13 y 15 años de edad y la declinación sexual entre
los 50 y 70 años aunque no es fenómeno fácil de precisar. En la mujer la pubertad se inicia
generalmente un poco antes, entre los 11 y 15 años, pero esta edad es bastante variable en
distintas partes del mundo; de inmediato aparecen fenómenos periódicos, a cargo sobre todo
de sus glándulas genitales, que sólo se interrumpen por algún embarazo o enfermedad; al
llegar a una edad cercana a los 50 años estos fenómenos desaparecen (menopausia) y con ellos
la etapa de fertilidad. Además algunos de los caracteres sexuales vuelven al estado prepuberal.
ANOMALIAS
Ya se ha indicado que es animal macho el que posee testículos y hembra el que posee ovarios,
y que ambas glándulas constituyen el carácter sexual primario.
La coexistencia en un mismo individuo de testículos y ovarios, llamada hermafrodismo
verdadero, es un hecho extraordinariamente raro en la especie humana, y del cual sólo existen
poco más de una decena de casos bien comprobados en la literatura médica mundial. Se
suelen observar con más frecuencia en los caracteres sexuales secundarios anomalías que
llevan a veces a la confusión de sexo (hermafrodismo falso o seudo hermafrodismo), pero el
diagnóstico real siempre debe basarse en la existencia de testículos u ovarios.
En muchas aves y batracios suelen observarse con cierta frecuencia las más curiosas
anomalías sexuales. En los faisanes se ve la coexistencia, en el mismo animal, del plumaje
masculino en una mitad del cuerpo y del femenino en la otra. Algunas gallinas al envejecer
toman ciertos caracteres de gallo, cantan, son combativas, persiguen a otras gallinas, incluso
pueden fecundarlas, y se
comprueba que junto al ovario izquierdo se ha desarrollado un
testículo que existía atrofiado.
DIFERENCIAS SEXUALES ENTRE EL HOMBRE Y LA MUJER
MANTENIMIENTO DE LOS CARACTERES SEXUALES: TESTICULO Y OVARIO
Los caracteres sexuales secundarios de uno y otro sexo son mantenidos directamente por un
conjunto de hormonas que provienen fundamentalmente del ovario o del testículo, y
accesoriamente de la suprarrenal o la tiroides. En cuanto a la hipófisis desempeña aquí
también un papel central que cumple principalmente por su acción sobre todas las otras
glándulas que acabamos de mencionar.
El testículo y el ovario son glándulas mixtas que producen una secreción que va al exterior
(secreción externa) y que es el gameto propio de cada sexo, espermatozoide para el testículo y
óvulo para el ovario, y otra secreción que, al pasar a la sangre (secreción interna) ejerce su
acción a distancia sobre órganos efecto
res determinados dando la característica propia de
cada sexo.
Fácil es demostrar el papel de la glándula de secreción interna mantenedora de los caracteres
sexuales del testículo y del ovario. La castración de los vertebrados, conocida y aprovechada
por el hombre desde tiempos remotos, no deja aparecer los caracteres sexuales si se
realiza
antes de la pubertad o borra alguno si es posterior. La inyección de las hormonas
correspondientes o el injerto de la glándula que falta, permiten la restitución de los caracteres
y hasta su exageración. Por ejemplo un gallo capón no canta, pierde los instintos sexuales, se le
atrofia la cresta, pero basta implantarle bajo la piel un testículo de otro gallo para que
reaparezcan los caracteres perdidos.
Acción de la castración y del injerto testicular sobre la cresta del gallo. A la izquierda, gallo
normal; al centro, gallo castrado; y a la derecha, gallo al que después de castrado se le hizo un
injerto testicular.
Ambas glándulas sexuales ejercen su acción por hormonas que fabrican y luego vuelcan en la
sangre. Estos últimos años han sido testigos de brillantes trabajos de fisiólogos y químicos que
han podido aislar de estas glándulas sus hormonas, conocer su fórmula química exacta y luego
prepararlas artificialmente en el laboratorio. De esta manera se ha llegado a comprobaciones
sorprendentes. Así el testículo prepara sustancias que inyectadas tienen acción masculina, y el
ovario otras que tienen acción femenina, pero además, y esto es muy curioso, cada glándula es
capaz de preparar en pequeño grado, hormonas del sexo opuesto, y cada hormona tiene
también y en pequeño grado, propiedades de las hormonas del otro sexo. Otro hecho notable
es que las hormonas sexuales conocidas hasta la fecha, así como las elaboradas por la corteza
suprarrenal, la vitamina D, etc., son de un parecido químico extraordinario y derivan de un
mismo antecesor químico; sus diferencias están en su acción y muy poco en su constitución.
En resumen, pequeñas diferencias en la constitución química pueden ir acompañadas por
grandes diferencias en la acción sobre el organismo.
REGULACION DE LAS GLANDULAS SEXUALES. PAPEL DE LA HIPOFISIS
El lóbulo anterior de la hipófisis estimula a las glándulas sexuales p o r medio de sustancias
llamadas gonadotrofinas hipofisarias, que produce y suprarrenal vierte en la sangre. Estas son
dos, una que actúa sobre la secreción externa madurando los gametos (óvulo y
espermatozoide), y otra desarrollando la secreción interna del testículo y del ovario, y, por su
intermedio, los caracteres sexuales secundarios.
La extirpación de la hipófisis, en un animal adulto, suprime la fuente de las gonadotrofinas
hipofisarias, se atrofian las glándulas sexuales y los caracteres sexuales específicos se borran o
atenúan. La inyección de gonadotrofinas, que son sustancias de naturaleza proteica, restituye
los caracteres, siempre que el ovario o testículo se hallen presentes.
Aparato genital de perras: a la izquierda, sin hipófisis; al centro, normal; a la derecha, sin
hipófisis inyectada con lóbulo anterior.
La secreción de gonadotrofina hipofisaria tiene variaciones y se conocen reguladores de la
misma de origen nervioso y hormonal. Los nerviosos han sido tratados al hablar de la hipófisis
y se explican por influencias que pasan del hipotálamo, en la base del cerebro, a la hipófisis.
Un corte de las vías nerviosas en este camino atrofia las glándulas sexuales por disminución
de la secreción de gonadotrofinas hipofisarias, y así, por ejemplo, deja de observarse en las
aves el enorme aumento de peso de los testículos que sigue a una intensa exposición del
animal a la luz. En cambio, la excitación eléctrica aumenta la secreción de gonadotrofinas y
trae fenómenos sexuales como es la puesta de óvulos en la coneja, la rata, etc.
Las influencias hormonales sobre la
producción hipofisaria de gonadotrofinas, hasta ahora
conocidas, son todas de orden inhibitorio. Así actúan las hormonas del testículo y el ovario y
también las de la tiroides y corteza suprarrenal. Por eso al castrar un animal y desaparecer el
freno que representan sus hormonas sexuales, aumenta el aflujo de gonadotrofinas
hipofisarias a la sangre.
En suma, el lóbulo anterior de la hipófisis estimula a las glándulas sexuales y por su
intermedio obra sobre los caracteres sexuales, regulando su actividad en base a estímulos
nerviosos que recibe el hipotálamo, y a inhibiciones hormonales que vienen del testículo o del
ovario, la tiroides o la suprarrenal.
Esquema del sistema endocrino que regula la función sexual.
Función sexual masculina. El testículo
El testículo, como el ovario, es una glándula mixta con una función
generativa, productora de espermatozoides o gameto masculino, y una función endocrina, que
consiste en la producción y entrega a la sangre de hormonas que mantienen los caracteres
sexuales secundarios masculinos, genitales y extragenitales. Aquí sólo se tratará la función
endocrina.
ANATOMIA E HISTOLOGIA
El aparato genital masculino está constituido por el testículo, órgano donde nacen los
espermatozoides, y un sistema de tubos que conducen a éstos al exterior. Como anexos a este
sistema se hallan varias glándulas cuyas secreciones se suman a la testicular. El aparato genital
masculino guarda íntimas relaciones anatómicas con el urinario y la parte final de sus vías es
común.
Arriba, esquema general del aparato sexual masculino; abajo, corte de un testículo en el cual se
pueden apreciar los lóbulos, los tubos seminíferos, el epidídimo y el conducto deferente.
Los testículos son dos órganos glandulares contenidos en las bolsas, fuera de las grandes
cavidades del cuerpo, lo que hace de ellos los órganos de más baja temperatura del organismo.
En el adulto tienen la forma y tamaño de un huevo de paloma y sus dimensiones son de 40 a
45 mm de largo; 25 mm de ancho y 30 mm de alto y cada uno pesa aproximadamente 20
gramos.
Una fuerte membrana, la albugínea, envuelve a cada testículo; de ésta parten tabiques que lo
dividen en unos 250 compartimientos cónicos, cuyos vértices confluyen en la parte posterior.
En cada compartimiento existen de 1 a 3 tubos, de 30 a 70 cm de largo por 150 a 250 micrones
de ancho, de curso muy tortuoso, llamados tubos seminíferos, los que sumados a los de los
demás compartimientos miden unos 250 metros. En las paredes de los tubos seminíferos hay
células que sufren un particular proceso de maduración que termina en la producción de
espermatozoides, que luego son vertidos en la luz del tubo seminífero. Esta maduración
solamente es completada a partir de los 14 años, edad que marca la pubertad en el varón, o sea
la iniciación de su vida sexual fértil y que continuará hasta los 50 ó 70 años.
El examen microscópico revela entre los tubos la existencia de vasos, nervios y células
reunidas en grupos, llamadas células intersticiales, a las que se cree productoras de la
secreción interna testicular.
Un largo y complejo sistema de tubos excretores conduce los espermatozoides desde los tubos
seminíferos hasta el exterior. Todos los tubos seminíferos convergen en el epidídimo, que es
una porción superpuesta al testículo como un casquete y dentro de la cual una serie de tubos
tortuosos dan origen al conducto deferente. Este tiene un espesor de 1 a 2 mm y un largo de
unos 40 cm. Penetra en el abdomen y desemboca en la uretra, cerca de su origen en la vejiga;
sin embargo, antes atraviesa la glándula prostática y da origen también a una dilatación en
forma de ampolla, la vesícula seminal. La uretra es un conducto mixto, génitourinario, por el
cual pasa la orina y la esperma; nace en la vejiga y se dirige hacia el pene, al que recorre en
toda su longitud, para abrirse al exterior en su extremidad libre.
Hay varias glándulas anexas al aparato genital que producen secreciones
que se vuelcan en
éste. La principal es
la próstata, situada en la pelvis, donde
es perforada por la uretra a su
salida
de la vejiga, como un tubo que perforara un tapón. La hormona sexual masculina
mantiene el tamaño y función normales de la próstata, la castración reduce notablemente su
tamaño. Después de los 40 ó 50 años de edad suele agrandarse en muchos sujetos, hasta
alcanzar a veces el tamaño de una naranja, y entonces ocasiona dificultades en la micción por
compresión de la uretra y otros serios trastornos que mejoran o curan con su extirpación.
HORMONAS MASCULINAS
El testículo fabrica una hormona, la testosterona, cuya acción es mantener los caracteres
sexuales masculinos, genitales y extragenitales. Si se la inyecta a un gallo castrado,
rápidamente le crecerá la cresta, volverá a cantar y perseguirá a las gallinas, y es éste el
procedimiento que habitualmente se sigue para investigar si existe testosterona o una
sustancia de acción masculina en un extracto. La testosterona sufre transformaciones en el
organismo y es eliminada por la orina. Se ha podido establecer su fórmula química que,
repetimos, es parecida a la de las hormonas fabricadas por el ovario, la corteza suprarrenal,
etc.
La corteza suprarrenal normalmente produce un poco de testosterona, y un exceso de la
misma en el caso de algunos tumores que atacan a la mujer.
REGULACION DE LA FUNCION TESTICULAR. LA HIPOFISIS
Al estudiar los caracteres sexuales se señaló la estrecha interdependencia hormonal entre el
testículo y el lóbulo anterior de la hipófisis, que en realidad constituyen una sola unidad
funcional.
El lóbulo anterior hipofisario segrega dos sustancias, las gonadotrofinas, que estimulan
selectivamente; una, la función generadora de espermatozoides, y otra, la endocrina. Estas
gonadotrofinas aumentan en el momento de la pubertad y a ellas se deben originariamente los
cambios que sufre el aparato sexual.
A su vez, el testículo por intermedio de la testosterona modera la hipófisis. Debe recordarse
que la hipófisis modifica su secreción de gonadotrofinas según estímulos que le llegan no sólo
del testículo, sino de la suprarrenal, de la tiroides y del sistema nervioso.
Esquema del sistema endocrino que regula la función sexual.
FUNCIONES DEL TESTICULO
Ya se dijo que son dos, la generativa y la endocrina. La generativa puede ser abolida por
simple ligadura de los conductos, que impide la producción de espermatozoides y deja
intactos los caracteres sexuales secundarios. Muy demostrativa es la separación de ambas
funciones, que se realiza aprovechando la existencia de las dos gonadotrofinas hipofisarias,
con acción estimulante sobre cada función. Si se extirpa la hipófisis faltan las dos
gonadotrofinas y se atrofia el testículo: no se producen espermatozoides y se borran los
caracteres sexuales secundarios. Inyectando ahora la que estimula la función generativa,
aparecerán los espermatozoides y el animal será fértil, pero desposeído de sus caracteres
sexuales secundarios; si por el contrario se inyecta la otra gonadotrofina los efectos son
contrarios.
A) CASTRACION. La castración o extirpación testicular doble, suprime bruscamente las dos
funciones. En los animales se la ejecuta desde la antigüedad con fines prácticos. En el hombre
se la observa por diversos motivos: religiosos, accidentales, guerreros, etc., y sus efectos
difieren un tanto según que el sujeto haya alcanzado o no la madurez sexual. La castración del
hombre adulto trae menos trastornos que la del joven, pues persisten muchos caracteres como
su barba, bigote, su tono de voz, su carácter y hasta sus instintos.
El castrado antes de la pubertad o eunuco, es tan característico en su aspecto exterior, que a
simple vista se lo puede reconocer. Suelen ser de gran estatura, con piernas largas y caderas
más anchas que los hombros; lampiños pero nunca calvos; con una voz de timbre especial que
los ha hecho formar parte de coros célebres; de carácter suave y de inteligencia normal. Sus
instintos sexuales no existen y sus órganos genitales muestran un aspecto infantil, condiciones
por las que eran empleados para cuidar los famosos serrallos de los sultanes.
Un hecho precoz y acentuado después de la castración pre o pospuberal, es la atrofia de las
glándulas anexas al aparato genital, como son la próstata y también las vesículas seminales,
cuyo estado normal depende directamente de la estimulación producida por las hormonas
masculinas.
B) RESTITUCION. Los síntomas de los castrados pueden ser corregidos por medio de la
inyección de hormonas masculinas o del injerto de testículo. El injerto suele realizarse con
éxito en los animales y también se ha empleado en el hombre con fines de rejuvenecimiento,
aunque lo que se ha conseguido es una temporaria excitación sexual, que ilusiona mucho a los
pacientes, mientras el injerto se reabsorbe, pues no prende como se creía.
C) HIPERFUNCION. Síntomas de hiperfunción endocrina testicular pueden ser obtenidos por
la inyección de hormonas testiculares en cantidad excesiva, o provocados por la aparición
espontánea de ciertos tumores que segregan en exceso a estas mismas hormonas. En ambos
casos hay un desarrollo supernormal de los caracteres sexuales secundarios en el varón o la
aparición de algunos de éstos en la mujer.
Cuando tumores testiculares de este tipo aparecen en niños provocan un desarrollo precoz de
la pubertad, crecen muy rápidamente, adquieren la forma y reciedumbre del hombre adulto y
madurez sexual; esto suele llegar a tal punto, que niños de 3 ó 4 años de edad pueden
confundirse con jóvenes de 15 ó 16 años. La resección del tumor cura los síntomas. Si tumores
con producción de hormonas masculinas aparecen en el ovario o la corteza suprarrenal de la
mujer, la masculinizan, le aparece barba
y bigote, adquiere gran fuerza muscular, carácter
masculino, etc.; la curación se obtiene al sacar el tumor.
En el hombre adulto normal el desarrollo de los caracteres sexuales es máximo; no es posible
una mayor respuesta a una mayor cantidad de hormona sexual.
Función sexual femenina. Ovario endocrino
El ovario es una glándula mixta que por una parte elabora el óvulo, para ser fecundado por el
espermatozoide y constituir el nuevo germen, y por otra fabrica productos hormonales que
pasan a la sangre y regulan la función sexual femenina en sus diversos aspectos. Es decir,
cumple una doble función: generativa y endocrina.
ANATOMIA DEL APARATO GENITAL DE LA MUJER
Comprende varios órganos situados en la parte inferior del abdomen (pelvis), destinados a
asegurar las funciones de fecundación y más tarde de la nutrición del feto. Además posee un
anexo, las glándulas mamarias, que después del nacimiento suministran la leche necesaria
para el hijo.
Los órganos genitales propiamente dichos son los ovarios, las trompas, el útero, la vagina y la
vulva.
Arriba, corte del aparato genital de la mujer; abajo, corte de un ovario mostrando los folículos
y el cuerpo amarillo así como el óvulo finalmente expulsado.
Los ovarios son dos, situados uno en la parte derecha e inferior de la pelvis y el otro en el
punto simétrico del lado izquierdo. En la mujer adulta su forma es la de un haba, y sus
dimensiones son de 2,5 a 5 cm de largo, 1,5 a 3 cm de ancho, 0,6 a 1,5 cm de espesor, y su peso
de 6 a 8 gramos. Su consistencia es resistente y su color grisáceo. En su superficie externa se
observan varias abolladuras, las que corresponden a los folículos en diverso grado de
evolución; alguna de éstas, de tamaño mayor que las restantes, puede pertenecer a un cuerpo
amarillo reciente o de embarazo.
El examen microscópico del ovario lo muestra constituido por un tejido de sostén, vasos y
nervios, y repartidas al azar miríadas de folículos.
El folículo ovárico es una unidad morfológica y funcional; existen en número de 400.000 por
ovario, pero en distintos grados de evolución. Comienza por ser una célula grande, que es el
futuro óvulo, rodeada por un conjunto de células foliculares; a medida que crece y madura el
óvulo, las células foliculares aumentan de número y terminan por dejar en
el centro del
folículo un espacio lleno de líquido; en la mujer adulta cada 28 días revienta un folículo y
expulsa al óvulo correspondiente, el que penetra en la trompa; el folículo vacío es invadido
por sangre, capilares sanguíneos y células, constituyendo el cuerpo amarillo, que es una
glándula endocrina. Los folículos que no llegan a transformarse en cuerpos amarillos
retrogradan y desaparecen (atresia folicular) y lo mismo sucede con el cuerpo amarillo, salvo
en caso de embarazo; entonces persiste unos 5 meses, constituyendo el cuerpo amarillo
verdadero. Se calcula que en la vida de una mujer sólo llegan a completar su evolución unos
300 ó 400 folículos.
Las trompas son dos tubos musculares, de 12 cm de longitud, por 1 cm de ancho, que se
extienden simétricamente desde el útero hasta el ovario correspondiente donde se ensanchan
en forma de pabellón. Su interior está lleno de repliegues y mojado por una secreción mucosa
que probablemente ayuda al huevo en su nutrición; las células contienen pequeñas
prolongaciones (cilias) dotadas de movimiento que facilitan la progresión del huevo hacia el
útero.
El útero es un órgano musculoso, hueco, situado en la parte baja de la pelvis, en comunicación,
arriba y a los costados con las trompas, y por debajo con la vagina. Tiene forma de pera
aplastada de adelante hacia atrás, con su vértice hacia abajo. Las dimensiones exteriores de
este órgano, en estado de reposo, son: longitud 7 cm, ancho 4 cm en las mujeres que no
tuvieron hijos y 1 cm más en las que tuvieron, y su peso de 50 a 60 gramos; durante el
embarazo aumenta extraordinariamente para contener el feto. La fuerte pared muscular se
halla revestida en su parte interna, mirando a la cavidad, por una membrana mucosa llamada
endometrio, que sufre cambios periódicos en su grosor y constitución durante cada fase de la
vida genital de la mujer. Sobre esto volveremos más adelante.
La vagina es un tubo muy elástico que se inserta alrededor del cuello del útero, es decir en su
polo pequeño, y se prolonga hacia abajo hasta desembocar en la vulva, formada por una serie
de repliegues mucosos por delante del ano.
En su conjunto el aparato genital femenino se halla formado por una serie de órganos
musculosos, elásticos y huecos, que comunican sus cavidades formando un solo conducto
desde los ovarios hasta el exterior. En este sistema se produce el acto de la cópula, la liberación
del óvulo y su encuentro con el espermatozoide (fecundación), el desarrollo del huevo
fecundado y luego su expulsión al exterior.
LOS CICLOS SEXUALES EN LA MUJER. MENSTRUACION
La vida genital de la mujer se inicia en la Argentina, por ejemplo, entre los 11 y los 15 años de
edad (t.m. 13 1/2) y persiste hasta los 45 ó 50 años. A la primera de estas fechas se la denomina
pubertad y coincide con el desarrollo de los órganos y caracteres sexuales, y a la segunda
menopausia o climaterio, y señala su declinación; una y otra van acompañadas de síntomas
generales muy marcados.
Durante el largo período que transcurre entre la pubertad y la menopausia hay una
periodicidad en las funciones genitales, que aumenta y disminuye en forma de ondas
regulares con tal que no la interrumpa un embarazo. A cada ondulación completa se la llama
ciclo y cada uno de éstos se compone de un período de actividad sexual, llamado estro o celo
en los animales porque en ellos es el único momento en que la hembra puede ser fecundada
por el macho, y un período de reposo sexual; como se verá luego, en la mujer las diferencias
no son tan absolutas. Los ciclos aparecen con un ritmo propio en cada especie animal: el
chimpancé, 36 días; la mujer, 28; la vaca, 28; la rata 4 a 6; la perra dos veces por año, etc.
Cada ciclo sexual comporta la aparición de modificaciones propias en cada
segmento del
aparato genital femenino, que en la mujer se estudian en dos fases sucesivas, folicular y del
cuerpo amarillo, gobernadas cada una por una hormona especial fabricada por la hipófisis.
Esquema que representa el ciclo menstrual femenino corriente, con las respectivas
modificaciones ováricas y del endometrio uterino
1º FASE, O FOLICULAR. Dura aproximadamente 14 días. Se observa en el ovario el
crecimiento y maduración de uno de sus folículos, que termina por estallar y liberar al óvulo
que penetra en la trompa. En las cercanías de este día se halla el período más fértil de la mujer.
Las hormonas ováricas (estrógenos) segregadas en esta fase alcanzan la trompa, el útero y la
vagina, donde producen modificaciones bien visibles al microscopio, y en especial el
progresivo aumento del espesor del endometrio o membrana interna del útero.
2º FASE, O DEL CUERPO AMARILLO. Dura como la anterior unos 14 días. Si el óvulo no es
fecundado, muere, pero mientras tanto el folículo que le dio origen se cierra y se llena de
células, que lo transforman en una verdadera glándula de secreción interna, el cuerpo
amarillo. La hormona segregada por el cuerpo amarillo completa el desarrollo del endometrio.
Si no hay fecundación, al final de la fase se rompen los vasos endometriales y escapa sangre
que arrastra las capas superficiales, por la vagina, al exterior. Este fenómeno, propio de la
mujer, se llama menstruación. Dura de 3 a 4 días, se repite cada 28 días y elimina de 20 a 200
cm3 de sangre, aunque la variabilidad de estas cifras suele ser grande incluso en los casos
normales. La menstruación marca no sólo el derrumbe de la mucosa uterina que a partir de allí
se rehace y reinicia un nuevo ciclo, sino también la inexistencia de la fecundación.
Es importante mencionar que a veces se producen todos los fenómenos descritos de un ciclo
menstrual completo, sin que haya puesta ovular. Estos ciclos anovulatorios se observan en
mujeres normales y sobre todo son frecuentes en las épocas de la pubertad y la menopausia.
Durante ellos la fecundación no es posible.
HORMONAS FEMENINAS
Las sustancias encargadas de mantener los caracteres sexuales femeninos, de ayudar y
mantener al embarazo, son varias y se clasifican en estrógenos y progesterona.
A) ESTROGENOS. El concepto de estrógeno es de orden muy general y comprende a toda
sustancia que inyectada a roedores castrados: rata, ratón, cobayo, etc. y por lo tanto en reposo
sexual, les provoca el estro o celo, es decir la actividad sexual. Se eligen los roedores para estos
ensayos por la comodidad de su manejo y la claridad con que muestran la producción del
estro. Dentro del concepto de estrógeno entran sustancias producidas por el organismo a las
que se llama estrógenos naturales, y otras de orígenes más diversos, que se llaman estrógenos
artificiales.
Los estrógenos naturales son hormonas producidas en el ovario y en la embarazada también
en la placenta, que pasan a la circulación y se encargan de mantener los caracteres sexuales
secundarios. A su acción estimulante se debe que el aparato genital femenino y las mamas,
tengan el aspecto y la estructura que vemos en la mujer adulta. Además toman parte activa,
por sus variaciones, en la producción de los ciclos periódicos que caracterizan la vida genital
de la mujer, y entre ellos el fenómeno de la menstruación. La prueba es que cuando la
producción de estrógenos se suprime extirpando el ovario, retroceden todos los caracteres
sexuales femeninos, disminuye grandemente el tamaño de sus órganos genitales y desaparece
la menstruación.
El ovario produce muy pocos estrógenos antes de la pubertad y después de la menopausia, en
el ínterin, durante la madurez sexual, esta secreción es abundante y sufre oscilaciones durante
los ciclos, a los que gobierna junto con la progesterona. Durante el embarazo su producción
aumenta, pues provienen también de la placenta. Los estrógenos son destruidos por el hígado,
y en parte se eliminan por la orina.
Los estrógenos naturales son varios: el estradiol, la estrona, etc. La fórmula química de estas
sustancias es muy parecida y tiene también grandes semejanzas con otras como son las
hormonas masculinas, las de la corteza suprarrenal, la vitamina D, etc. Se los puede preparar
por síntesis química.
Los estrógenos artificiales no son hormonas en el sentido que hemos dado a esta palabra. Se
extraen de diversas fuentes (plantas, alquitrán de hulla, etc.) o se fabrican en el laboratorio. Se
conocen varios: el dietilestilbestrol, el hexestrol, etc. Su fórmula química difiere
profundamente de las naturales, pero tienen iguales efectos fisiológicos. Son muy utilizados en
Medicina por su bajo costo y el hecho de ser activos si se toman por vía bucal.
B) PROGESTERONA. La progesterona es la hormona del cuerpo amarillo del ovario, y en la
embarazada también de la placenta. Debe considerarse como la hormona destinada a
mantener el embarazo y su papel es complementario de los estrógenos naturales. El cuerpo
amarillo la produce y vierte en la sangre, desde la pubertad hasta la menopausia, con
variaciones periódicas caracterizadas por su aparición e incremento en las dos semanas
previas a la menstruación, y por su desaparición brusca con esta última. También aumenta en
forma notable y persistente durante el embarazo.
Conviene hacer resaltar su particular acción sobre el endometrio, al que prepara para anidar al
huevo y mantener al feto y, además, sobre la mama, que llega así a un estado preparatorio
para segregar leche. La progesterona es eliminada por la orina. Se la puede fabricar en el
laboratorio por síntesis.
REGULACION DE LA FUNCION OVARICA
El ovario produce estrógenos y progesterona, ésta en el cuerpo amarillo. Ambas secreciones se
hallan en íntima dependencia del lóbulo anterior de la hipófisis, que por medio de las dos
gonadotrofinas estimula independientemente la producción de cada una. A su vez los
estrógenos y la progesterona moderan la producción de las gonadotrofinas hipofisarias, con lo
que se establece un doble juego hormonal de regulación mutua. Además actúan sobre la
hipófisis estímulos que llegan por el sistema nervioso o de otras glándulas como la tiroides y
las suprarrenales.
Resulta de lo dicho que la secreción de hormonas por el ovario es, fundamentalmente, el
resultado de un mecanismo endocrino hipofisoovárico.
FUNCIONES DEL OVARIO
Se dijo en la parte general que el ovario tiene dos funciones: la generativa, o sea la producción
del óvulo que pasa a las trompas para ser fecundado, y la endocrina, o sea la producción de
hormonas que vierte en la sangre. Gracias a estas últimas mantiene los caracteres sexuales
secundarios de la mujer, el embarazo y prepara las mamas para la secreción de leche.
La extirpación de un solo ovario es ampliamente compensada por el otro, pero la castración
doble trae síntomas profundos de insuficiencia ovárica que difieren si es ejecutada antes o
después de la pubertad. La prepuberal no deja aparecer los caracteres sexuales secundarios:
los órganos genitales y las mamas quedan infantiles, no aparecen las variaciones cíclicas, etc.
La castración pospuberal, reduce inmediata y grandemente el tamaño de los órganos genitales
y de las mamas, desaparecen los ciclos y se observan serios trastornos, tanto nerviosos como
del carácter y de la conducta.
Los trastornos producidos por la castración son curados por el injerto ovárico o la
administración bien regulada de los estrógenos y la progesterona, al punto que se pueden
reproducir hasta los ciclos de la mujer normal.
Si durante el curso de un embarazo se destruye en la coneja, el cuerpo amarillo, cesa la
producción de progesterona y el embarazo se interrumpe (Fraenkel, 1901). Esto no se observa
en otras especies animales porque la placenta suple la producción de progesterona.
Fecundación
Por fecundación se designa la formación del huevo, comienzo del nuevo ser, por la fusión del
espermatozoide o gameto masculino con el óvulo o gameto femenino. La necesidad del
encuentro de ambos gametos fue demostrada primeramente por Spallanzani, en 1787, y
comprobada luego numerosas veces.
DESCRIPCION
El momento óptimo para la fecundación de la mujer se halla a mediados del ciclo, es decir más
o menos 14 días antes de la menstruación, pues es cuando se rompe el folículo maduro y se
libera el óvulo que en seguida cae, entra en la trompa por su pabellón y circula lentamente en
dirección al útero. En la trompa se produce la fecundación por encuentro con el
espermatozoide, que viene ascendiendo desde el fondo de la vagina, donde fuera depositado
durante
el acto sexual, a través del útero. La ascensión del espermatozoide se debe a
movimientos propios debidos a su cola, a los que se agrega en la trompa la ayuda que le
prestan los movimientos de las pestañas de las células superficiales de la misma. La cabeza de
uno solo de los millones de espermatozoides presentes penetra en el óvulo y se fusionan los
núcleos, en tanto que su cola se pierde. El óvulo así fecundado baja al útero y anida en uno de
los repliegues de la mucosa uterina donde se segmenta y continúa su desarrollo ulterior.
Esquema de la fecundación del óvulo por el espermatozoide. A) Penetración de un
espermatozoide; B,C) división del óvulo. Abajo, espermatozoide visto con gran aumento.
DETERMINACION DEL SEXO
El sexo del futuro ser queda decidido en el mismo momento del encuentro de los gametos por
la calidad del espermatozoide y no por la del óvulo. En efecto, hay espermatozoides de dos
tipos según los cromosomas de su núcleo; uno de ellos dará lugar a varones y el otro a
mujeres. Por lo tanto, el hecho de que nazca un varón o una mujer es fruto en cierto modo
aparentemente de la casualidad, aunque la naturaleza, por razones que todavía no se conocen,
mantiene parecidas proporciones de nacimientos de varones y mujeres.
DIFERENCIACION SEXUAL
El embrión no presenta en los primeros tiempos caracteres que permitan reconocer el sexo a
que pertenece, pues es bisexuado, es decir que en él se hallan desarrollados los esbozos de
ambas glándulas sexuales. Más adelante se establece la diferenciación y sigue su desarrollo un
sexo solamente. Este origen embrionario común explica algunas de las anormalidades sexuales
observadas en el adulto.
Gestación
El huevo que anidó en la mucosa uterina o endometrio, crece, se transforma, y al mismo
tiempo se producen modificaciones importantes en el organismo materno. Las modificaciones
fetales y las maternas serán consideradas por separado, pero señalaremos antes, que la
duración del embarazo es bastante fija para cada especie animal:
Mujer: 280 días
Vaca: 280 días
Yegua: 340 días
Chimpancé: 236 días
Oveja: 150 días
Perra: 60 días
Coneja: 30 días
Rata: 22 días
MODIFICACIONES FISIOLOGICAS MATERNAS
Desde la fecundación, y acentuándose a medida que progresa el embarazo, van apareciendo
modificaciones en los órganos genitales y también fuera de ellos.
A) MODIFICACIONES GENITALES. Todo el aparato sexual fememino sufre modificaciones a
medida que progresa el embarazo: la vagina, las trompas, las mamas, pero donde adquieren
un carácter de particular intensidad es en el útero.
a) Uterinas. Durante la gravidez en la mujer el útero crece tanto que su peso, de 50 g al
comienzo del embarazo, alcanza, al final, 1.000 g sin su contenido, y su capacidad interior
aumenta en igual período de sus 2 a 5 cm3 hasta 5.000 ó 7.000 cm3. Este cambio extraordinario
se debe en primer lugar al desarrollo de la capa muscular, cuyas fibras aumentan en largo
unas 10 veces y también en número, y accesoriamente al endometrio que aumenta de espesor
y vascularización.
b) Placenta. En la parte del endometrio donde anida el huevo se producen modificaciones muy
especiales que llevan a la constitución de un órgano perfectamente individualizado, la
placenta. En la mujer la placenta está situada en la cara interna de la pared del útero, tiene la
forma de una torta y su tamaño, al fin del embarazo, es de unos 20 cm de largo por 2 cm de
ancho y su peso de 500 gramos. De sus dos caras, una se apoya sobre la pared del útero y la
otra mira al feto; de esta última parte el cordón umbilical, con una longitud aproximada de 50
a 60 cm de largo y un espesor de 1 a 2 cm, que es el encargado, por venas y arterias que corren
por su interior, de llevar y traer sangre al feto.
La placenta, órgano intermediario entre la madre y el hijo, es la encargada de realizar los
intercambios de nutrición y respiratorios entre ambos, y en su constitución intervienen partes
maternas y fetales. El huevo, desde que se implanta en el endometrio, emite un sinnúmero de
vellosidades en forma de dedo de guante, que penetran en los anchos lagos sanguíneos que
forman los vasos maternos y flotan en ellos. En el interior de esas vellosidades se desarrollan,
poco a poco, arterias y venas que se comunican con las fetales, de tal manera que la sangre de
la madre y la del hijo, aunque vecinas al nivel de las vellosidades, se hallan separadas por la
pared de las mismas como por un filtro, y nunca se comunican directamente. La disposición
señalada corresponde a la mujer, pues en otras especies animales es distinta.
A través de la pared de las vellosidades pasan, de la madre al hijo, el agua y las sales disueltas,
oxígeno, vitaminas, hormonas, etc., y vuelven en sentido contrario los residuos fetales. La
pared sirve para interceptar el paso a microbios y partículas de cierto tamaño, y desempeña así
un papel de protección.
La placenta es también un importante órgano de secreción interna; sobre este papel se insistirá
más adelante.
B) MODIFICACIONES EXTRAGENITALES. El organismo materno presenta numerosas
modificaciones como consecuencia del embarazo, las que alcanzan a casi todas sus funciones.
Su mención detallada llevaría mucho espacio.
En la piel aparecen manchas, las que se acentúan al nivel de los pezones y en la cara, donde en
algunas mujeres constituyen la llamada máscara del embarazo.
Además aumenta la retención de muchas sustancias que deben ser utilizadas para la nutrición
del feto, como las proteínas, el calcio, el fósforo, el hierro, el yodo, etc. El agua es retenida en
gran cantidad y es uno de los factores de aumento del volumen sanguíneo.
Las combustiones son mayores en los últimos meses del embarazo por la suma de las fetales a
las maternas, aumentando así el desprendimiento de calor por la superficie de la madre.
C) PRODUCCION DE HORMONAS: DIAGNOSTICO DEL EMBARAZO. El embarazo se
caracteriza porque gran cantidad de hormonas aparecen en la sangre y orina de la mujer;
provenientes todas ellas del lóbulo anterior de la hipófisis, del ovario, y de la placenta.
El lóbulo anterior de la hipófisis aumenta su producción de gonadotrofinas que pasan a la
sangre y escapan por la orina.
El ovario aumenta su producción de estrógenos y sobre todo de progesterona, esta última
proveniente del cuerpo amarillo, que persiste hasta el 4º ó 5º mes del embarazo.
La placenta es la fuente principal de los estrógenos que escapan por la orina; da poca cantidad
de progesterona y además segrega en los primeros tiempos del embarazo una hormona propia
que también se descubre en la orina, la gonadotrofina coriónica.
La identificación de esta hormona en la orina de una mujer permite diagnosticar un embarazo,
a partir de los 15 días de la fecundación, con casi un 100% de seguridad. Esto es sumamente
importante, porque ningún otro medio conocido puede hacerlo con tal precocidad. La
conocida reacción de Friedman consiste en inyectar a una coneja orina de la presunta
embarazada y observar el animal 48 horas después; si los ovarios tienen unos puntos
hemorrágicos (folículos) el diagnóstico es positivo.
Reacción de Friedman para diagnosticar embarazo. Negativa, y positiva con hemorragia
visibles
Galli Mainini ha demostrado en la Argentina que igual resultado puede obtenerse inyectando
la orina a un sapo macho, el que responde con descarga de espermatozoides en la orina a las 2
ó 3 horas. Ambas reacciones de, embarazo y otras, se fundan en la presencia de gonadotrofina
coriónica, que es la sustancia causante de las modificaciones señaladas, en la orina.
D) PARTO. Llegado a su término el desarrollo del feto, la madre comienza a sentir dolores,
luego se rompen las membranas que envuelven al feto, escapa el líquido que contienen y por
una serie de etapas perfectamente sincronizadas se produce su expulsión del claustro materno.
El trabajo de parto dura aproximadamente de 12 a 16 horas en las primíparas, y 6 a 10 en las
multíparas, y es debido a contracciones dolorosas de las fibras musculares del útero, cada vez
más fuertes y cercanas, que van empujando su contenido hacia el exterior. Una vez expulsado
el feto y al cabo de un tiempo variable de 1 a 2 horas, se produce el alumbramiento o
eliminación de la placenta. En seguida el útero se contrae cerrando así sus vasos que sangran,
y desaparece la cavidad dejada por el feto y sus anexos. En los días siguientes se produce
lentamente la expulsión del endometrio y todo el aparato genital pasa a la situación de reposo
y reparación.
DESARROLLO DEL FETO
El huevo fecundado se desarrolla rápidamente y necesita alrededor de 9 meses para completar
su evolución, habiendo pasado antes por el estado que se denomina de embrión hasta el tercer
mes, y por el de feto de allí en adelante. Las profundas transformaciones sufridas durante este
lapso son motivo de estudio por una ciencia especial, la Embriología. Aquí sólo se tratarán
algunos puntos de fisiología fetal.
La circulación del feto tiene dos profundas diferencias con la del adulto, causadas por la
existencia de un circuito placentario y la falta de circulación pulmonar, ya que el feto no utiliza
sus pulmones. Por el circuito placentario recibe el feto los materiales alimenticios, hormonales,
el oxígeno, etc., que le da la madre, y a él lleva sus materias de desecho, sin ponerse nunca en
contacto directo ambas sangres, que se hallan se paradas por la membrana de las vellosidades
coriales. Dos arterias van por el cordón umbilical al feto y una vena vuelve por él. Al nacer,
esta circulación se interrumpe y se inicia la pulmonar, por la que se harán en adelante los
intercambios de los gases respiratorios.
La interrupción brusca, al nacer, de la circulación placentaria y el estado de asfixia inmediata
son las principales causas estimulantes de la primera respiración, que pondrá así en actividad
la circulación pulmonar. El frío que sufre el feto al pasar al medio externo y los toques sobre la
piel son factores estimulantes que coadyuvan para iniciar la respiración.
El peso del recién nacido oscila generalmente entre 3 y 31/4 kg. En la Argentina diversas
estadísticas arrojan valores medios para los varones de 3,4 kg y las niñas de 3,5 kg.
Secreción mamaria y leche
Todos los mamíferos segregan leche, que es el alimento esencial del recién nacido y uno de los
principales de las poblaciones humanas.
La madre tiene la obligación natural y moral de dar su leche al hijo, con lo que le suministra
los materiales indispensables para su desarrollo y su defensa contra las infecciones. Con razón
se dice, "que la leche de la madre pertenece al hijo".
DESARROLLO DE LAS MAMAS Y SUS FASES DE SECRECION
A) ANATOMIA. En la mujer existen dos glándulas mamarias con un mamelón en su centro,
situadas simétricamente a ambos lados del pecho. En el vértice del mamelón hay de 15 a 25
orificios de 0,7 mm, de los que parten tubos de 2 a 4,5 mm de luz, que se dividen y subdividen
hasta terminar en una extremidad ciega, ensanchada, constituida por las células secretoras de
la leche. El conjunto de racimos que desembocan en
cada orificio constituye un lóbulo
mamario. Los lóbulos se disponen alrededor del mamelón como los radios de una rueda. Cada
lóbulo constituye, en verdad, una glándula mamaria independiente, separada de las demás
por grasa y tejidos de sostén.
B) FASES DE DESARROLLO. Las mamas permanecen infantiles hasta la edad de la pubertad,
en que sufren un brusco desarrollo; luego sufren pasajeros aumentos de tamaño en la primera
mitad del embarazo y durante la lactancia. Por el contrario, se produce una reducción
definitiva después de la menopausia o de la castración.
C) FASES DE SECRECION. La producción de leche por las mamas sólo existe
en
determinados períodos de la vida de la mujer coincidentes con el desarrollo anatómico.
En el recién nacido suele haber durante unos días una ligera secreción llamada leche de bruja,
y que es debida al pasaje de hormonas de la madre al feto; una secreción similar se observa en
la madre al fin del embarazo y en seguida del parto, hasta tanto se produce la subida de la
leche. Iniciada ésta, 2 ó 3 días después del parto, llega a su máximo 6 ó 9 meses después de él,
y suele extinguirse generalmente al año y medio.
Un estímulo muy importante para mantener la secreción láctea es la succión que ejerce el hijo.
MECANISMO HORMONAL DEL DESARROLLO DE LA MAMA Y LA SECRECION
LACTEA
REGULACION DE LA SECRECION LACTEA
El desarrollo de la glándula mamaria y la secreción de leche son dos fenómenos gobernados
por factores diferentes. Es vulgar la observación de que una mama bien desarrollada no
segregue leche, pero el desarrollo previo es un paso favorable para la producción láctea.
El desarrollo de la mama en la pubertad, el embarazo o la lactancia, se debe principalmente a
los estrógenos y progesterona segregados por el ovario (y la placenta al final del embarazo).
Los estrógenos desarrollan los conductos y la progesterona las células del fondo de ellos, que
luego segregará la leche. La prueba del papel estimulante del ovario es dada por la atrofia
mamaria que sigue a la castración.
Para que una mama así desarrollada segregue leche, es necesaria la acción estimulante de la
prolactina hipofisaria. Esta hormona aislada pura del lóbulo anterior de la hipófisis por
Riddle, aparece después del parto, pasa a la sangre y llega a la mama. Coadyuva con esta
acción otra nerviosa y estimulante ejercida por el niño al succionar del mamelón.
LECHE
La leche, o producto específico de secreción de las glándulas mamarias, es un líquido cuya
producción oscila entre 600 a 1.000 cm3 por día. Extraída del organismo o por acción del jugo
gástrico coagula, es decir pasa del estado líquido al sólido.
A) ORIGEN. Las sustancias que la componen provienen de la sangre que llega a la glándula,
pero ésta no actúa como un filtro inerte sino que deja pasar unos materiales y fabrica otros,
concentrando cada uno en proporciones casi fijas. Algunas de las sustancias ingeridas por la
madre pueden llegar al hijo por esta vía y causarle trastornos, por ello es bueno saber que tal
es el caso del alcohol, del opio, la belladona, algunos purgantes, etc.
B) COMPOSICION QUIMICA. La leche tiene casi un 90% de agua y su densidad es poco
mayor que la de ésta (de 1,028 a 1,034).
En su composición entran sustancias orgánicas y minerales. Entre las primeras hay varias
proteínas como el caseinógeno, que por acción del cuajo del estómago se transformará en
caseína; la lactalbúmina y la lactoglobulina, análogas a las de la sangre. La lactoglobulina
contiene las sustancias maternas defensivas de los gérmenes. El azúcar es la lactosa. Las grasas
son varias; a ellas debe la leche su opacidad, su color y su sabor y se hallan flotando en forma
de tenues gotitas que al subir forman la nata. Las sustancias minerales son numerosas: potasio,
calcio, sodio, fósforo, cloro, notándose únicamente la relativa escasez de hierro.
A las sustancias anteriores hay que agregar la presencia de vitaminas A, B, C en gran cantidad,
y de muy poca vitamina D.
C) VALOR NUTRITIVO. La leche es un alimento de primera clase, esencial para el niño, y uno
de los más importantes para el adulto. Para el recién nacido es el mejor alimento. Las
poblaciones que toman mucha leche son fuertes y las que no pueden beberla débiles. El
hombre la utiliza en estado natural, o se vale de derivados que son muy apreciados, como la
crema, el queso, la manteca. Las principales deficiencias son su pobreza en hierro y en
vitamina D.
Cada especie animal tiene un tipo de leche propia y adaptado a sus crías, a las cuales
suministran los elementos que les son indispensables. Mal puede entonces una especie animal
ser alimentada con leche perteneciente a otra, que no tiene ni igual composición química, ni los
elementos protectores contra las mismas enfermedades. En este terreno, por ejemplo, la leche
de mujer difiere de la de vaca, como se observa en el cuadro, no solamente porque es más rica
en lactosa y grasa y pobre en proteínas y sales, sino que en la de vaca están presentes los
anticuerpos contra las enfermedades del ternero. Además, la digestibilidad es distinta y el
coágulo de la leche de mujer se hace en pequeñas partículas y el de vaca en una masa grande.
Composición química de la leche en gramos por 100 cm3
Los órganos de la locomoción
Las funciones de respiración, di gestión, circulación, secreción interna y excreción tienen por
finalidad conservar constante el medio interno. Veamos ahora qué dispositivos tiene el
organismo para ajustarse como un todo al medio externo.
En los capítulos que siguen describiremos el esqueleto, que es el armazón del ser viviente y
tiende a mantener su forma y posición, al sistema muscular que mueve al cuerpo y a sus
diferentes segmentos.
DESCRIPCION DEL ESQUELETO
El esqueleto del hombre está construido por unos 200 huesos cuya enumeración escaparía a los
fines de este tratado (lámina III). Los huesos están constituidos por laminillas dispuestas en
forma muy característica, y formadas por un tejido compacto y rico en sales de calcio. Si las
laminillas se disponen apretadamente el tejido óseo es compacto, si son ralas, esponjoso. En los
espacios del tejido esponjoso se encuentra una sustancia blanda, que interviene en la
formación de los glóbulos rojos, la medula ósea.
PRINCIPALES HUESTOS DEL ESQUELETO
Los huesos pueden tener diversas formas generales: cortos, planos, largos. Los huesos largos
tienen por lo general una cavidad central en la parte media angosta y larga (la diáfisis) y dos
extremidades abultadas o epífisis de tejido esponjoso.
Esquema que representa un corte longitudinal de una tibia, hueso largo del cuerpo humano.
Entre la epífisis y la diáfisis se encuentra en los niños un disco de tejido cartilaginoso (cartílago
de conjunción), lugar donde se forma hueso nuevo permitiendo el crecimiento. Cuando el
crecimiento termina, el disco es reemplazado por tejido óseo.
Esencialmente los huesos están dispuestos alrededor de un eje central, longitudinal, la
columna, formada por 33 ó 34 vértebras, huesos compuestos de un cuerpo anterior y un arco
que limitan un agujero grande, el agujero vertebral. La superposición de estos agujeros forma
un conjunto que aloja a la medula espinal.
La primera vértebra, el atlas, se articula con el cráneo constituido por huesos planos, algunos
con prolongaciones, que delimitan una cavidad, la cavidad craneal, donde se aloja el sistema
nervioso superior: cerebro, cerebelo y bulbo.
Al cráneo se unen por delante y abajo los huesos de la cara.
El atlas y las 6 vértebras que lo siguen hacia abajo constituyen las vértebras cervicales. Luego
vienen 12 vértebras, llamadas dorsales, a las que se articulan de cada lado 12 huesos largos y
curvos: las costillas, formando una especie de, jaula más angosta por arriba que por abajo,
llamada tórax. Las costillas se vinculan por delante con el esternón.
Las clavículas y los omóplatos son huesos vinculados al tórax que forman la cintura escapular.
A ellos se unen los miembros superiores, constituidos por el húmero, hueso del brazo; el radio
y el cúbito, huesos del antebrazo, y luego los huesos de la muñeca y de la mano.
Unido a las últimas vértebras que forman un bloque rígido (el sacro) se halla a cada lado un
hueso, el coxal o ilíaco, formando un arco que constituye la cintura pelviana; a ella se articulan
miembros inferiores constituidos por el fémur, hueso del muslo; la tibia y el peroné, huesos de
la pierna, y los huesos del tobillo y del pie.
LAS ARTICULACIONES
Los distintos huesos del cuerpo están unidos entre sí por articulaciones, rígidas o movibles.
Entre las rígidas mencionaremos las articulaciones de los huesos del cráneo; entre las
movibles, hay algunas de movimientos limitados, como la articulación entre dos vértebras, o
de movimientos amplios como la articulación del hombro.
Las articulaciones muy movibles son de varios tipos: tipo bisagra, en la que el movimiento
sólo puede hacerse en el eje transversal, por ejemplo la articulación de los dedos de la mano;
tipo rueda, en la que el movimiento sólo se puede hacer en el eje longitudinal, por ejemplo el
componente de la articulación del codo que permite al antebrazo rotar como si fuera un
destornillador del brazo; tipo bola y esfera, como las articulaciones del hombro y de la cadera
que permiten movimientos en tres ejes.
LOS MUSCULOS
Los músculos estriados o esqueléticos. cuyo número en el cuerpo es alrededor de 500,
producen por su contracción movimientos cuya naturaleza dependerá de sus puntos de
inserción. La mayoría de los músculos se insertan en sus dos extremos en huesos diferentes.
Sin embargo, hay algunos que no tienen ninguna inserción ósea (músculos de los labios,
esfínter anal, etc.), y otros que se insertan por un extremo en el hueso y por el otro en la piel
(músculos de la mímica) o en bandas fibrosas.
He aquí los principales músculos posteriores del hombre: 1) Trapecio; 2) Deltoides; 3)
Infraespinoso; 4) Romboides; 51 Redondo menor; 6) Tríceps braquial; 7) Dorsal ancho; 8)
Glúteo mediano; 9) Anióneo; 10) Cubital posterior; 11) Glúteo mayor; 12) Extensos común de
los dedos; 13) Semitendinoso; 14) Bíceps crural; 15) Recto interno; 16) Semimembranoso; 17)
Sartorio; 18) Gemelo interno; 19) Gemelo externo; 20) Soleo; 21) Flexor largo del dedo gordo;
22) Flexor largo de los dedos; 23) Tendón de Aquiles. Los músculos que se representan son de
los llamados estriados o esqueléticos de fibras alargadas.
Entre los músculos que mueven las articulaciones mencionaremos el bíceps y el tríceps, flexor
y extensor del antebrazo respectivamente.
Articulación del codo y los músculos tríceps y bíceps. En negro, huesos y tendones; en blanco,
los músculos.
Pero ha de tenerse en cuenta que casi nunca actúa un músculo aisladamente; siempre se
contraen simultáneamente grupos de músculos. Cuando flexionamos el antebrazo no sólo se
contrae el bíceps sino otros músculos sinergistas. También la contracción de los músculos que
contribuyen a flexionar el antebrazo supone la relajación o inhibición de otros músculos que,
de otra manera, se opondrían a ese movimiento: los antagonistas.
ESTRUCTURA DE LOS MUSCULOS. La
propiedad de producir movimiento no es exclusiva
del músculo. Existen en el organismo otras células capaces de moverse, como los glóbulos
blancos de la sangre y las células que poseen cilias vibrátiles. Pero sin duda, de todos los
tejidos del cuerpo, son los músculos los que más desarrollada tienen esta actividad. Existen
tres tipos de músculos.
Esquema que representa los tres tipos de estructura del músculo.
1) el músculo liso, que se encuentra en las paredes del tubo digestivo, del aparato urinario y de
los vasos sanguíneos. La contracción de este tipo de músculos es poco vigorosa y muy lenta y
no depende de la voluntad. En efecto, los músculos lisos están inervados por el sistema
nervioso autónomo que, como se verá más adelante, es involuntario. Están formados por
células cuyas fibras son cortas y no tienen estriación
transversal; 2) el músculo estriado,
llamado así por estar formado por células alargadas, llenas de fibrillas, en las que se observan
bandas transversales claras y oscuras, lo que les da un aspecto muy característico. Los
músculos esqueléticos están constituidos por tejido muscular estriado y su contracción es
provocada por estímulos voluntarios que provienen del sistema nervioso central; 3) el músculo
cardíaco, que se asemeja en su estructura al músculo esquelético. Sin embargo, sus estriaciones
son más próximas, y sus células están unidas entre sí formando una extensa red. Las
contracciones del músculo cardíaco se producen debido a estímulos provenientes del corazón
mismo (automatismo).
PROPIEDADES DEL MUSCULO ESTRIADO. a) Excitabilidad. Se denomina así a la propiedad
del músculo de responder a un estímulo directo con una contracción. Se puede hacer contraer
al músculo por medio de estímulos mecánicos, térmicos, químicos y eléctricos. Pero en
condiciones fisiológicas el músculo es estimulado exclusivamente a través de sus nervios. Un
músculo privado de nervios es incapaz de contraerse.
Como se verá más adelante, cada nervio motor está constituido por numerosas fibras que al
llegar a la cercanía del músculo se dividen en ramas (hasta 120), terminando cada rama en una
fibra muscular mediante un órgano especial: la placa motriz.
Terminaciones de una fibra nerviosa en una fibra muscular.
Al llegar el impulso nervioso a la placa motriz, se produce un retardo durante
el cual se
transmite el proceso de excitación del nervio al músculo. En este proceso interviene como
intermediario químico la acetilcolina, liberada en las terminaciones nerviosas, en la placa
motriz.
b) Contractilidad. El músculo excitado se contrae: sus fibras se acortan y ensanchan sin
cambiar su volumen total. Si no se opone resistencia al acortamiento del músculo no se
modifica la tensión de éste y la contracción se denomina isotánica. Si por el contrario se opone
una resistencia tal que se impida el acortamiento del músculo, durante su contracción se
desarrolla una gran tensión en el músculo, sin modificación de su tamaño; es la contracción
isométrica.
La estimulación simple de un nervio motor provoca una contracción aislada: después de un
breve período latente el músculo se acorta hasta un máximo y luego se relaja. Es la sacudida
simple.
Gráfico de una sacudida simple y de una contracción tetánica. La línea superior representa el
gráfico obtenido conectando el músculo con un registrador. La línea media representa la
aplicación de los estímulos.
Durante unas 5 milésimas de segundo después de un estímulo eficaz el músculo está en
período refractario; es decir, es incapaz de responder a un segundo estímulo. Si se envía una
serie de estímulos a intervalos algo mayores que la duración del período refractario, se obtiene
una contracción sostenida, mucho más fuerte que la producida por un estímulo aislado. Este
tipo de contracción se denomina tetánica, y es el tipo habitual de contracción de los músculos
esqueléticos.
c) Efecto de la intensidad de los estímulos. La aplicación de estímulos aislados de muy
pequeña intensidad a un nervio motor no provocará respuesta en el músculo. Los estímulos
son subliminales. Si se aumenta gradualmente la intensidad del estímulo llegará un momento
en que el músculo responde con una contracción muy débil (umbral); luego junto con la
intensidad de los estímulos aumentará gradualmente la fuerza y amplitud de la contracción
hasta llegar a un límite (estímulo máximo), más allá del cual, por intensos que sean los
estímulos (estímulos supramáximos), la fuerza de la contracción no variará. Esta gradación de
la fuerza de la contracción no se debe a una gradación de la fuerza con que se contraen
individualmente las fibras musculares. Estas responden como el corazón según la ley del todo
o nada. Lo que sucede es que en las contracciones débiles por estímulos apenas
supraliminares, sólo unas pocas fibras se contraen, y a medida que aumentan los estímulos es
mayor el número de fibras que entran en contracción hasta que cuando el estímulo es máximo,
todas ellas se contraen.
d) Tono muscular. Los músculos están en el organismo en un estado permanente de
contracción parcial, que
consiste en un tétanos suave, producido por series continuas de
impulsos nerviosos que afectan a pequeños grupos de fibras musculares alternadamente.
Como veremos, el tono muscular es un reflejo, el reflejo miotático de Sherrington.
e) Fenómenos químicos de la contracción muscular. El análisis químico de los músculos, antes
y después de contraerse, revela que en el músculo activo se encuentra menor cantidad de
glucógeno (un hidrato de carbono de elevado peso molecular), de fosfatos orgánicos, y mayor
cantidad de ácido láctico, anhídrido carbónico, fosfatos inorgánicos, y que se consume
oxígeno. Los prolijos estudios realizados revelan la participación de muchas otras sustancias
en el proceso mediante el cual se transforma la energía química en mecánica, durante el
proceso de contracción. Nos limitaremos a referir esquemáticamente los tres tipos de
reacciones que se llevan a cabo.
El estímulo nervioso provoca la transformación explosiva de fosfatos orgánicos (fosfocratina y
adenilpirofosfato) en fosfatos inorgánicos y otros compuestos orgánicos con liberación de
energía. Si no se restituyeran los fosfatos orgánicos pronto se agotarían las fuentes de energía
para la contracción. Pero la resíntesis de esas sustancias requiere un aporte de energía química.
Esta es proporcionada por la transformación de glucógeno en ácido láctico, que se efectúa con
gran rapidez. También la cantidad de glucógeno es limitada en el músculo, y además la
acumulación de ácido láctico en él provoca trastornos que se traducen por la fatiga. Recién en
esta etapa interviene el oxígeno aportado por la sangre. Las cuatro quintas partes del ácido
láctico se convierten nuevamente en glucógeno. Pero este proceso de resíntesis del glucógeno
también requiere energía, y la fuente de energía, en este caso, es la combustión de la quinta
parte restante del ácido láctico que, combinada con el oxígeno, da como producto de
combustión anhídrido carbónico, agua y energía.
f) Fenómenos térmicos y eléctricos de la actividad muscular. No toda la energía química
liberada durante la contracción muscular se convierte en energía mecánica. Lo mismo que en
cualquier motor de combustión una buena parte de la energía se libera en forma de calor. La
relación entre la energía transformada en trabajo y energía total movilizada para producirlo es
lo que se denomina: rendimiento. Las mediciones realizadas en el músculo indican que sólo un
20 ó 30% de la energía utilizada se convierte en trabajo mecánico. En este sentido el músculo es
un motor de buen rendimiento, pues los motores a vapor tienen un rendimiento del 15% y los
de gasolina del 18%. Los motores Diesel en cambio tienen un rendimiento mayor, que puede
llegar al 35%.
El calor desarrollado por los músculos no es como en los motores puramente un producto de
desecho. En efecto, sirve para mantener la temperatura del cuerpo alrededor del óptimo
necesario para el buen funcionamiento del organismo. La contracción muscular es uno de los
métodos que utiliza el organismo para mantener constante su temperatura. Uno de los
mecanismos fundamentales para la lucha contra el frío en el hombre es el escalofrío con
temblor. La cantidad de calor que esta actividad muscular produce es considerable, puesto que
los músculos esqueléticos constituyen cerca del 50% del peso del cuerpo.
La llegada del estímulo nervioso al músculo hace que se produzca en éste una variación
eléctrica. La porción de la fibra muscular excitada se hace eléctricamente negativa con respecto
a las partes próximas. Esta variación eléctrica, que puede registrarse mediante dispositivos
adecuados, se propaga a toda la fibra muscular, pero no pasa a las fibras vecinas.
Sistema nervioso
El organismo está compuesto por infinidad de células que forman tejidos que se agrupan a su
vez en órganos
o sistemas. Cada célula puede considerarse una unidad en sí misma. Sin
embargo, su agrupación en tejidos para formar el cuerpo del hombre no es una colección
anárquica. El hombre, lo mismo que cualquier animal, es un individuo multicelular, pero que
actúa y responde como una unidad. Sus múltiples funciones están coordinadas y reguladas
por mecanismos diversos que componen un todo de las distintas partes, es decir integran la
unidad funcional.
La integración funcional se efectúa por dos medios: humorales y nerviosos. Entre los agentes
humorales de integración mencionaremos: a) las sustancias nutritivas; b) las secreciones de las
glándulas endocrinas; c) productos del metabolismo, como el anhídrido carbónico: d)
sustancias químicas producidas en el organismo (acetilcolina, simpatina, histamina).
El sistema nervioso es, sin embargo, por su rapidez y precisión de acción y su complejidad el
factor principal del mecanismo de integración. Sin él, el organismo sólo podría adaptarse
pobremente al medio externo.
DESCRIPCION DEL SISTEMA NERVIOSO
Anatómicamente el sistema nervioso del hombre puede describirse como un tubo de forma
variable de cuya parte superior, el encéfalo, que ocupa la cavidad craneana, parten 12 pares de
nervios craneales, y de cuya parte inferior, la medula espinal, que ocupa el canal vertebral,
parten 31 pares de nervios espinales.
Un corte sagital de la cabeza permite reconocer en el encéfalo varias porciones distintas: 1) el
bulbo (mielencéfalo) que es la continuación hacia arriba de la medula espinal; 2) la
protuberancia anular y el cerebelo (metencéfalo); 3) el mesencéfalo, constituido por los
tubérculos cuadrigéminos y los pedúnculos cerebrales; 4) el diencéfalo (tálamo, hipotálamo,
etc.) y por último, 5) el telencéfalo, que comprende los ganglios de la base y la corteza cerebral.
La parte hueca del tubo, que constituye en la medula el canal central, modifica su forma en el
encéfalo: al nivel del bulbo y protuberancia se ensancha formando el cuarto ventrículo; luego
vuelve a estrecharse al nivel del mesencéfalo formando el conducto de Silvio, el cual al nivel
del diencéfalo desemboca en el tercer ventrículo, comunicado lateralmente con los ventrículos
cerebrales del telencéfalo.
Representación esquemática de un corte sagital del encéfalo y de la medula espinal. En el
encéfalo se pueden observar todas las partes que lo componen.
LA NEURONA: UNIDAD ANATOMICA DEL SISTEMA NERVIOSO
El sistema nervioso está constituido por células de gran excitabilidad, vinculadas entre ellas de
tal manera que constituyen vías conductoras de estímulos. Estas células son las neuronas, que,
aun cuando presentan ciertas diferencias estructurales según la porción del sistema nervioso
que se considere, tienen varias características anatómicas y fisiológicas comunes.
Esquema de los dos tipos de neurona.
El cuerpo celular provisto de un núcleo muestra en su citoplasma finos hilos que forman una
red y que se introducen en las prolongaciones celulares: son las neurofibrillas. Cada célula
nerviosa emite prolongaciones que pueden clasificarse en dos tipos: axones y dendritas. Cada
célula emite por lo menos un axón, raras veces dos o más. Se caracterizan estas prolongaciones
por su grosor uniforme, diámetro pequeño, pocas ramificaciones y, en general, por su gran
longitud. Hay axones de más de 1 metro de largo. Cada célula emite por lo general varias
dendritas, que se caracterizan por ramificarse abundantemente a corta distancia de su célula
de origen, por su contorno irregular, calibre variable y breve longitud. Cada neurona
constituye una unidad anatómica. Su axón y sus dendritas no
se unen directamente con los
axones y dendritas de otras neuronas, sino que toman contacto anatómico y se ponen en
relación funcional, por lo que se denomina sinapsis. En ella el axón de una neurona se pone en
comunicación con el cuerpo o las dendritas de otra neurona. Nunca se establecen conexiones
funcionales entre las dendritas de una neurona y el cuerpo celular o las dendritas de otra.
El impulso nervioso se propaga a través de la sinapsis mediante la liberación, al nivel de las
terminaciones del axón de la primera neurona, de una sustancia, la acetilcolina, que excita a las
dendritas o cuerpo de la neurona con la que se ha puesto en contacto. iniciando en ésta otro
impulso nervioso.
Las fibras nerviosas son axones cubiertos o no por una vaina de sustancia grasa, aisladora, la
mielina. La vaina de mielina sufre interrupciones a intervalos regulares (nódulos de Ranvier).
Estructura de la fibra nerviosa.
Las fibras nerviosas pueden ser de cuatro clases: a) fibras mielínicas, recubiertos de una
membrana fina (neurilema). Constituyen los nervios periféricos. b) fibras mielínicas sin
neurilema, que se hallan en el sistema nervioso central. c) fibras amielínicas, especialmente
numerosas en el sistema simpático, y d) axones desnudos sin mielina y sin neurilema,
abundantes en la sustancia gris del cerebro y de la medula.
La vaina de mielina es lo que da el
aspecto blancuzco de los nervios periféricos y de la
sustancia blanca del sistema nervioso central; la ausencia de mielina hace que las fibras
nerviosas tengan un color gris. En el sistema nervioso central se distinguen claramente dos
tipos de estructura: la sustancia gris y la sustancia blanca. La primera está formada por células
nerviosas con sus prolongaciones desnudas. La segunda por fibras mielínicas.
LA CONDUCCION DEL ESTIMULO NERVIOSO. El estado de excitación de un músculo se
revela por su acortamiento o su tensión y el de una glándula por su secreción, pero en cambio
ningún signo visible nos revela el pasaje de un estímulo a través de un nervio.
La aplicación de un estímulo a un punto del nervio produce en éste ciertos cambios (excitación
local) que se propagan a lo largo de la fibra como la llama por un reguero de pólvora
(excitación propagada). La transmisión del impulso nervioso es un proceso que va
acompañado de fenómenos metabólicos (aumento del consumo de oxígeno, producción de
calor, acumulación de metabolitos) y eléctricos.
Al excitar un nervio con una corriente eléctrica u otro estímulo eficaz se produce en el sitio
excitado un potencial eléctrico. Si éste alcanza cierto nivel crítico (umbral) se produce un
aumento brusco de potencial, que se puede registrar con métodos que captan y amplifican las
variaciones eléctricas. En los trazados, este aumento de potencial se registra como un pico o
aguja que se propaga a lo largo del nervio. La amplitud del pico es máxima una vez que el
estímulo ha llegado al umbral. También el nervio sigue, pues, la ley del todo o nada. Se puede
medir la velocidad de conducción del impulso nervioso colocando en dos puntos alejados del
nervio electrodos que capten la variación eléctrica propagada, a su paso por cada uno de los
puntos, y conociendo la distancia que los separa. Erlanger y Gasser, cuyos estudios sobre la
fisiología nerviosa les valieron el Premio Nobel, clasificaron las fibras nerviosas según su
velocidad de conducción en fibras A, B y C, dividiéndose el grupo A en 5 subgrupos. Las
fibras de mayor velocidad de conducción son las del grupo A, que son fibras con mielina,
motrices y sensoriales. Su diámetro alcanza hasta 20 micrones, y su velocidad de conducción
hasta 120 m por segundo. El grupo B está constituido por fibras mielínicas de pequeño calibre
pertenecientes al sistema simpático, con velocidades de conducción de 3 a 14 m por segundo.
El grupo C está formado por fibras muy finas amielínicas, cuya velocidad de conducción no
pasa de 2 m por segundo.
Como se ve, existe una relación directa entre velocidad de conducción y diámetro de la fibra
nerviosa.
Los nervios periféricos se componen de fibras de distinto diámetro y por consiguiente de
distinta velocidad de conducción.
FUNCION TROFICA DE LA NEURONA. La
neurona es también una unidad trófica.
Cualquier parte de la neurona que se separe de la porción celular que contiene el núcleo,
muere, mientras que la parte que permanece en conexión con el núcleo sobrevive. La
degeneración del fragmento distal del axón se extiende a sus más finas ramificaciones, pero no
pasa a las sinapsis y no afecta a otras neuronas vecinas.
Después de seccionar un axón la célula reacciona: se producen cambios en el citoplasma y en
el núcleo y, como consecuencia de estos cambios, el extremo cortado del axón comienza a dar
brotes que crecen en varias direcciones; pero si el extremo distal degenerado se halla, a poca
distancia, penetran de preferencia en él y siguen por los tubos formados por las vainas de las
fibras que quedan como residuo del proceso degenerativo, hasta las últimas terminaciones del
net, vio degenerado. Los restos del nervio cortado sirven, pues, de tutores a los brotes que
crecen del extremo seccionado.
VIAS DE CONDUCCION. Un estímulo que parte de la periferia hacia los centros superiores, o
que parte de éstos hacia la periferia, sigue vías bien definidas en las que participan más de una
y a veces varias neuronas, formando una especie de cadena que puede esquematizarse así para
una vía motora:
Y para una vía sensitiva:
Los impulsos que parten de los centros hacia la periferia se denominan eferentes. Los que
provienen de la periferia y van hacia los centros se denominan aferentes.
La conducción a través de una fibra nerviosa, sea ésta una dendrita o un axón, se puede
realizar en todo sentido, como se transmite en cualquier dirección la corriente eléctrica en un
alambre conductor. Pero en la sinapsis la conducción se efectúa en un solo sentido: el impulso
nervioso sólo puede pasar a través de la sinapsis en la dirección del axón de una neurona a
dendrita o cuerpo de la otra, y así sucesivamente; nunca en sentido inverso. Las sinapsis
actúan, pues, como válvulas.
Al nivel de la sinapsis el estímulo sufre un pequeño retardo e incluso puede ser bloqueado.
EL REFLEJO: UNIDAD FUNCIONAL DEL SISTEMA NERVIOSO
Cuando un estímulo actúa sobre un receptor adecuado, y al pasar por las vías aferentes se
traduce en una respuesta debido a la llegada, a un efector, de un estímulo que viaja por vías
eferentes, se dice que se ha producido un reflejo. El reflejo es la unidad funcional del sistema
nervioso.
Un arco reflejo, en su forma más simple, está constituido por: a) el receptor; b) la neurona
aferente; c) la sinapsis; la neurona eferente y e) el efector.
Esquema que representa al arco reflejo más simple
Los receptores son formaciones especializadas para recibir y ser excitadas por estímulos que
provienen del exterior (órganos de la visión, del tacto, del oído), o del interior del organismo
(vísceras, músculos, etc.).
De los receptores nacen fibras nerviosas aferentes que constituyen las prolongaciones
dendríticas de neuronas sensitivas. Por ellas van hacia la célula nerviosa impulsos originados
en los receptores.
El axón corto de esta neurona aferente se pone en contacto a través de una sinapsis con la
dendrita de una neurona motriz. Esta neurona eferente transmite por su axón el impulso
nervioso hacia el efector. Se denomina así a órganos que responden al estímulo nervioso
modificando su actividad (músculo, glándula, corazón, etc.).
Esta descripción del arco reflejo es excesivamente esquemática. Por lo general existen una o
más neuronas intercaladas entre la neurona aferente y la eferente (neuronas internunciales).
Además la neurona aferente hace sinapsis con más de una neurona internuncial o eferente, de
modo que al ser excitado el receptor se modifica la actividad de muchas neuronas eferentes,
pudiendo entrar en juego varios efectores. Por ejemplo, la excitación de un receptor o de un
nervio aferente no hace contraer o relajar por vía refleja a un músculo determinado, sino a un
conjunto de músculos, que ejecutan por su acción combinada un movimiento.
A) CARACTERISTICAS DE LOS REFLEJOS. a) Período latente. Es el intervalo de tiempo entre
la aplicación de un estimulo al receptor y la respuesta motriz reflejada. Parte del retardo se
debe al tiempo necesario para que el impulso nervioso recorra las vías periféricas aferente y
eferente. Si se resta este tiempo periférico, que puede medirse, del período latente del reflejo,
se tiene el tiempo central del reflejo o retardo central. Este retardo central se debe
principalmente al retardo sináptico, y es tanto mayor cuanto mayor sea el número de neuronas
internunciales, y por consiguiente de sinapsis, que intervengan en el arco reflejo.
b) Suma y facilitación. La estimulación de un nervio aferente provoca en la célula eferente del
arco reflejo un estado de excitación; si el estímulo ha sobrepasado el umbral, la célula eferente
descarga un impulso. Si el estímulo es subliminal (por debajo del umbral) la célula eferente no
descarga un impulso, pero el estado de excitación producido por la llegada del estímulo
persiste durante cierto tiempo, y puede sumarse con otros estados de excitación provoca dos
por impulsos subliminales que llegan a la célula eferente sucesiva (suma temporal) o
simultáneamente por otras vías (suma espacial). De manera que la reiteración de estímulos
subliminales o la llegada simultánea de varios estímulos, cada uno de los cuales es incapaz de
por sí de determinar una respuesta, se suman y provocan la descarga de la célula eferente.
Si el estado de excitación persistente en la célula eferente ha sido provocado por un estímulo
que ha provocado respuesta, se habla de facilitación.
c) Posdescarga. En un nervio periférico estimulado cesa todo efecto al dejar de estimularlo. En
cambio en un arco reflejo la respuesta del órgano efector continúa hasta varios segundos
después de haber cesado el estímulo del órgano receptor. Esto es debido, probablemente, a la
persistencia en las neuronas internunciales del efecto producido por la estimulación de los
aferentes.
d) Inhibición. Inervación recíproca. Se denomina inhibición la supresión de la actividad de un
efector por acción nerviosa. Un ejemplo claro de inhibición nerviosa es el paro cardíaco
producido por la excitación del vago. En este caso la inhibición es periférica: el impulso
nervioso inhibe la actividad del efector y sabemos que lo hace por la liberación de un
intermediario químico: la acetilcolina. Otro tipo de inhibición lo constituye el principio de
Inervación recíproca. La excitación del nervio sensitivo de un miembro produce la flexión del
mismo (reflejo de flexión) y la extensión del miembro del lado opuesto (reflejo de extensión
cruzada). Pero al mismo tiempo que se contraen los músculos flexores del miembro excitado y
se contraen los músculos extensores del lado opuesto, se relajan (se inhiben) los músculos
antagonistas: extensores en el miembro excitado y flexores en el del lado opuesto. La
inhibición en este caso no se debe a una inhibición periférica, pues no existen nervios
relajadores de los músculos. Se debe a una inhibición central, que tiene por resultado la
disminución de los impulsos que normalmente envían las neuronas motoras a los músculos
que se relajan en el reflejo de flexión.
B) TEORIA HUMORAL DE LA TRANSMISION SINAPTICA.
Todas las características de los
reflejos que hemos señalado pueden explicarse admitiendo que en la transmisión de la
excitación nerviosa a través de la sinapsis intervienen mediadores químicos del tipo de la
acetilcolina.
El período latente prolongado se debería a que en cada sinapsis del reflejo, se requiere cierto
tiempo para la elaboración del intermediario químico.
El efecto de suma se explicaría por la desaparición relativamente lenta del intermediario
químico al nivel de la sinapsis, lo que permitiría su acumulación por aportes sucesivos de
cantidades insuficientes hasta sobrepasar la cantidad necesaria para provocar la descarga. La
posdescarga también se debería a la persistencia del intermediario.
En cuanto a la inhibición central, podría explicarse por la desaparición más rápida del
intermediario excitante, o por la existencia de otra sustancia de propiedades inhibitorias.
C) TEORIA ELECTRICA DE LA TRANSMISION SINAPTICA.
Si bien existen argumentos
experimentales importantes en favor de la teoría humoral, también existen otros que permiten
explicar la transmisión sináptica como un fenómeno puramente eléctrico. Cuando el potencial
eléctrico en pico llega a la sinapsis, después de haber recorrido el axón, puede excitar a la
neurona siguiente en la misma forma que lo haría una descarga eléctrica aplicada en ese
punto.
D) CLASIFICACION DE LOS REFLEJOS. Los reflejos pueden clasificarse de varias maneras
según el punto de vista que se considere.
a) Según el tiempo de aparición. Se denominan reflejos innatos o incondicionales los que
existen desde el nacimiento. Los que se adquieren en el curso de la vida son los reflejos
adquiridos o condicionados. Se mencionó un ejemplo de éstos al hablar de secreción salival.
b) Según el número de segmentos que intervengan. Los reflejos cuyo arco ocupa un solo
segmento medular se llaman reflejos segmentarios. Aquellos cuyo arco ocupa varios
segmentos se denominan reflejos intersegmentarios. Un ejemplo de éstos es el reflejo del
rascado. La excitación de una zona del dorso y flanco de un perro provoca la flexión y
extensión rítmicamente alternada de la pata posterior del mismo lado.
c) Según el tipo de receptores. Se denominan reflejos enteroceptivos los que tienen su origen
en las vísceras; reflejos exteroceptivos son los que tienen sus receptores en la piel y en los
órganos de los sentidos y son provocados por estímulos externos; y reflejos propioceptivos,
aquellos cuyos receptores se hallan en los músculos, tendones, articulaciones y laberinto.
d) Según el nivel del sistema nervioso en que se efectúen. Así hay reflejos medulares, bulbares,
protuberanciales, etc.
E) ALGUNOS TIPOS DE REFLEJOS.
Se denomina reflejos a respuestas características a
estímulos específicos.
a) Un grupo importante de los reflejos tipos lo constituyen los reflejos de defensa, dirigidos a
remover estímulos nocivos o a alejar al organismo de dichos estímulos. El más simple es el
reflejo de flexión, que consiste en la flexión de un miembro al cual se ha aplicado un estímulo
doloroso. La percepción del dolor no es factor indispensable pues el reflejo se produce en
animales descerebrados.
b) Otro grupo lo constituyen los reflejos posturales, denominados así porque gracias a ellos se
mantienen la postura y el equilibrio del cuerpo. El más típico es el reflejo de estiramiento, o sea
la contracción de un músculo en respuesta al estiramiento de sus fibras. Por ejemplo el reflejo
rotuliano: la percusión del tendón de los músculos extensores de la pierna provoca su
estiramiento. Existen en el tendón y en el músculo receptores propioceptivos, sensibles a la
distensión que, por vía refleja, determinan la contracción de los extensores de la pierna.
Estos reflejos de estiramiento tienen gran importancia, pues de ellos depende la
semicontracción permanente de los músculos que hemos mencionado al hablar del tono
muscular, y también depende de ellos, en parte principalísima, el mantenimiento de la
postura.
LA MEDULA ESPINAL
La medula espinal es un tubo de sustancia nerviosa alojado en el canal vertebral y que emite
simétricamente a cada lado 31 pares de nervios.
En un corte transversal se ve que está formada por una masa central de sustancia gris, que
adopta la forma de una H rodeada por todos lados de sustancia blanca. La sustancia gris está
formada por células nerviosas y fibras amielínicas, y la sustancia blanca por fibras nerviosas
con vaina de mielina. Las fibras de la sustancia blanca son: fibras aferentes que provienen de
los receptores periféricos y hacen estación en las células de la sustancia gris; fibras aferentes
que llevan impulsos de todos los segmentos de la medula hacia los centros encefálicos,
disponiéndose en haces longitudinales cuya localización es conocida; fibras de las neuronas
internunciales de los arcos reflejos (fibras de asociación); fibras eferentes que provienen de los
centros superiores y hacen estación en las células de la sustancia gris de la medula; y fibras
eferentes que parten de las células de la sustancia gris de la medula hacia la periferia.
En la parte relativa a sensibilidad superficial y profunda se estudiarán las vías y centros
nerviosos de la sensibilidad por lo que no insistiremos aquí. Sólo describiremos las vías
motrices.
Las fibras nerviosas que provienen de las células de la corteza en la zona motriz después de
atravesar la cápsula interna, los pedúnculos cerebrales y la protuberancia, llegan al bulbo. Allí
la mayoría de las fibras se cruzan formando en la medula el haz lateral córticoespinal o haz
piramidal cruzado, terminando en las células del asta anterior, de donde parten las fibras
eferentes para los músculos. La gran mayoría de las fibras que no se han cruzado al nivel del
bulbo lo hacen más abajo, de manera que la estimulación de la corteza motriz del lado
derecho, provoca movimientos en el lado izquierdo del cuerpo y viceversa. Por lo mismo un
derrame cerebral del lado derecho, que comprime y destruye las fibras motrices al nivel del
cerebro, provoca una hemiplejia, una parálisis del lado izquierdo del cuerpo.
Esquema del curso de las fibras nerviosas que provienen de la zona motriz de la corteza
cerebral.
A) LOS NERVIOS RAQUIDEOS.
De la medula espinal parten simétricamente 31 pares de
nervios: son los nervios raquídeos.
Cada nervio tiene dos raíces: una anterior o ventral, que lleva fibras eferentes, y una posterior
o dorsal que lleva fibras aferentes. Estas fibras aferentes después de abandonar el nervio y
antes de entrar en la medula hacen estación en un ganglio, donde están situados los cuerpos
celulares de dichas fibras.
Cada uno de los nervios raquídeos se distribuye por un sector del cuerpo. Así por ejemplo los
músculos del brazo están inervados por fibras motrices provenientes del 59 par cervical.
Igualmente se ha podido limitar las zonas periféricas de sensibilidad correspondientes a cada
par raquídeo.
B) SECCION DE LA MEDULA ESPINAL. La sección completa de la medula espinal es seguida
por la pérdida total y permanente de todo movimiento voluntario de los músculos cuya
inervación motora procede de segmentos de la medula situados por debajo del nivel de
sección, así como de la sensibilidad. Si la sección se ha realizado entre el bulbo y la medula, se
tiene lo que se llama un animal decapitado o espinal. En este caso se interrumpen las vías
eferentes que gobiernan los músculos respiratorios y el animal muere si no se le hace
respiración artificial. Una sección por debajo del primer segmento dorsal provocará la parálisis
de los músculos del abdomen y del miembro inferior, sin afectar a los músculos respiratorios
ni a los del miembro superior.
Cuando se produce la sección completa de la medula por una bala u otro tipo de lesión aguda
en el hombre, siente éste la sensación de que ha desaparecido parte de su cuerpo. Se produce
luego un estado de shock (shock espinal) en el cual no sólo desaparecen las sensaciones y la
motilidad voluntaria en las regiones correspondientes al extremo seccionado de la medula,
sino que también desaparecen los reflejos medulares de esa zona. Estos reflejos no tardan sin
embargo en reaparecer y con ellos reaparece, en cierto grado, el tono muscular y la acción
refleja de los esfínteres vesical y anal, lo que permite la evacuación refleja de la vejiga y del
recto.
C) SISTEMA NERVIOSO AUTONOMO. El sistema nervioso regula y coordina las funciones
de las vísceras mediante una división del mismo que tiene ciertas características especiales
que justifican su descripción aparte. El sistema se compone de vías aferentes, centros y vías
eferentes, por donde se establecen reflejos que regulan las funciones de las vísceras y aparatos
del organismo. Las vías aferentes nacen en las distintas vísceras (aparatos digestivo,
circulatorio, respiratorio, urinario, genital, endocrino, etc.). Los impulsos que provienen de
estas vías no causan por lo general sensaciones conscientes.
Las vías eferentes que tienen una disposición anatómica particular que luego describiremos,
tampoco están bajo la dependencia de la voluntad ni de la conciencia.
De manera que esta división del sistema nervioso no depende de la voluntad (sistema
nervioso involuntario), tiene poca conexión con el resto del sistema nervioso central
(autónomo), y regula la vida orgánica o vegetativa (sistema nervioso vegetativo).
Todas las vísceras reciben por lo general una doble inervación proveniente del sistema
autónomo: una excitante y otra inhibidora.
A) Disposición anatómica. Las características anatómicas de este sistema son las siguientes: a)
el cuerpo celular de la última neurona eferente se halla fuera del neuroeje y a veces en la
vecindad del órgano inervado; b) sus fibras nerviosas son amielínicas y de tiempo de
conducción largo.
Puede dividirse el sistema autónomo en dos sistemas según el origen de sus fibras eferentes: el
sistema simpático (ortosimpático o toracolumbar), y el sistema parasimpático (cráneosacro).
b) Sistema simpático. Las vías eferentes nacen en células de la parte antero lateral de la
sustancia gris de la medula espinal.
Nacimiento de las fibras nerviosas del sistema simpático.
Las fibras nerviosas emergen de la medula con las raíces anteriores de los nervios raquídeos
dorsales y dos o tres primeros lumbares, y van a terminar en ganglios que forman una cadena
a lo largo de la columna vertebral. La fibra que va de la medula al ganglio se denomina
preganglionar. De las células nerviosas del ganglio parten las fibras posganglionares que,
después de un recorrido más o menos largo, van a terminar en su elector correspondiente. El
sistema simpático tiene diversas acciones que pueden ser excitantes o inhibidoras.
Esquema que representa la inervación de algunas vísceras por el sistema autónomo.
c) Sistema parasimpático. Las vías eferentes nacen en neuronas: 1) situadas en el mesencéfalo,
protuberancia y bulbo, y salen del neuroeje con los pares craneanos III, VII, IX y X; 2) situadas
en la medula sacra y que salen con las raíces anteriores sacras.
Estas fibras preganglionares siguen un trayecto a veces muy largo y van a parar a ganglios
situados por lo general en la vecindad o aun en el interior mismo del órgano efector.
El sistema parasimpático tiene diversas acciones que pueden ser excitantes o inhibidoras. El
par X, neumogástrico o vago, es el nervio parasimpático más importante, pues lleva libras
eferentes preganglionares que van a terminar en ganglios situados en el corazón, el hilio del
pulmón, el plexo esolágico y los plexos del abdomen superior y del estómago o intestino.
d) Acción recíproca del simpático y del parasimpático. Casi todos los órganos de la vida
vegetativa reciben doble inervación: simpática y parasimpática, siendo por lo general de
acción opuesta. Así, por ejemplo, en el corazón el simpático es acelerador, el parasimpático es
inhibidor; en el intestino el parasimpático es excitador, el simpático inhibidor; en la pupila el
simpático es dilatador, el parasimpático constrictor; en los vasos sanguíneos, el simpático es
constrictor y el parasimpático es dilatador. La acción general del simpático es la de movilizar
la energía del organismo en una forma amplia y se manifiesta sobre todo en los estados de
emergencia, cuando se debe realizar un esfuerzo; por ejemplo en la lucha, en las emociones
intensas: miedo, ira, etc. El parasimpático, en cambio, tiene una acción más localizada, y sus
efectos tienden a acumular o restituir energía.
e) Transmisión humoral. Existen numerosas pruebas de que la acción de los nervios del
sistema autónomo se efectúa mediante sustancias químicas liberadas al nivel de sus
terminaciones nerviosas en los efectores.
El simpático libera simpatina, sustancia pariente de la adrenalina; y el parasimpático libera
acetilcolina. En los órganos periféricos existen sustancias que oxidan a la adrenalina, con lo
que desaparece rápidamente su acción. También existe una enzima, la colinesterasa, que
destruye a la acetilcolina.
Al nivel de la sinapsis ganglionar de ambos sistemas se desprende acetilcolina, que es la
sustancia responsable de la transmisión sináptica.
f) Acción de las drogas. Las drogas que tienen una acción similar a la del simpático se
denominan simpático miméticas. El ejemplo típico es la adrenalina. Las que suprimen la
acción del simpático se denominan simpaticolíticas; figuran en este grupo la ergotamina, la
iohimbina y otras sustancias sintéticas.
Las drogas que tienen una acción similar a la del parasimpático se denominan
parasimpaticomiméticas. La acetilcolina y otros derivados de la colina, y la pilocarpina son las
más conocidas. Las que suprimen la acción del parasimpático se denominan
parasimpaticolíticas y la droga tipo es la atropina, principio activo de la belladona.
D) CENTROS REFLE JOS SUPERIORES.
La
medula espinal se continúa hacia arriba con el
bulbo. En éste varía algo la disposición de la sustancia gris, que se interrumpe para dejar paso
a fibras nerviosas que se entrecruzan. Del bulbo parten los pares craneales del VI al XII, y en él
se encuentran varios centros reflejos importantes que ya han sido mencionados en otros
capítulos. Recordemos entre ellos el centro respiratorio, el cardiomoderador, el vasomotor, el
centro de la deglución y el del vómito.
En la sustancia gris de la protuberancia hacen estación fibras que provienen de la corteza. De
las células nerviosas de la protuberancia parten fibras nerviosas que van al hemisferio
cerebeloso del lado opuesto.
En el mesencéfalo están situados ciertos centros de la visión y de la audición.
En el hipotálamo está situado el centro regulador de la temperatura corporal y otros que
intervienen en la regulación de la presión arterial, del sueño y del metabolismo El hipotálamo
es, pues, el centro integrador de las reacciones viscerales más complejas. El tálamo es una
estación de las vías sensoriales (ver capítulo sensaciones).
NERVIOS CRANEALES
Del encéfalo se desprenden doce pares de nervios denominados pares craneales. Son éstos, de
adelante hacia atrás:
I. Nervio olfatorio, que conduce las sensaciones olfativas desde la mucosa pituitaria de la nariz
hasta el bulbo olfatorio del cerebro.
II. Nervio óptico, que conduce las sensaciones visuales desde la retina hasta los centros
corticales de la visión.
III. Nervio motor ocular común, que nace del pedúnculo cerebral y transmite impulsos
motores a músculos de la órbita.
IV. Nervio patético, que nace del pedúnculo cerebral y transmite impulsos motores al músculo
oblicuo mayor del ojo.
V. Nervio trigémino, nervio mixto motor y sensitivo, que nace de la protuberancia y conduce
fibras motrices a los músculos masticadores. Su raíz sensitiva conduce hacia el bulbo fibras
sensitivas provenientes de la cara (piel de la cara, dientes) y mitad anterior de la cabeza.
VI. Nervio motor ocular externo, que nace de la parte inferior de la protuberancia y conduce
fibras motoras para el músculo recto externo del ojo.
VII. Nervio facial, nace en el bulbo y envía fibras motoras a los músculos de la cara y fibras
secretorias a las glándulas salivales. También lleva al bulbo sensaciones del gusto originadas
en la parte anterior de la lengua.
VIII. Nervio auditivo, que lleva a los centros las sensaciones auditivas por su rama codear, y
por su rama vestibular los estímulos originados en los canales semicirculares y demás
formaciones del oído interno que constituyen, en conjunto, el laberinto.
IX. Nervio glosofaríngeo, nervio mixto, que del bulbo conduce impulsos motores a los
músculos de la faringe y estímulos secretorios a la glándula parótida, y lleva hacia los centros
fibras sensitivas de la parte posterior de la lengua y de la mucosa faríngea.
X. Nervio vago, nervio mixto, como el anterior, que nace en el bulbo. Por él van fibras
eferentes al corazón, esófago, estómago e intestino delgado, músculos de la laringe, glándulas
secretorias del estómago, y vienen fibras aferentes de los pulmones, de la mucosa de la laringe,
de la aorta, del estómago, etc.
XI. Nervio espinal, que inerva los músculos de la cintura escapular (hombro).
XII. Nervio hipogloso, que inerva los músculos de la lengua.
POSTURA Y MOVIMIENTO
Los músculos de nuestro cuerpo no cuelgan libres de sus inserciones óseas; aun en reposo
están en estado de semicontracción permanente (tono muscular). Ya hemos mencionado el
mecanismo reflejo del tono muscular, que es
,
uno de los factores principales en el
mantenimiento de la postura del cuerpo.
El tono muscular es, como ya se sabe; un reflejo propioceptivo originado en: el mismo músculo
por su estiramiento Pero este reflejo medular es modificado) por influencias que provienen de
los centros superiores. De estas influencias las hay que favorecen el tono proveniente de la
corteza cerebral y de los núcleos vestibulares. El shock espinal, que sucede a la sección
completa de la, medula se debe a la supresión de esos
,
impulsos facilitadores. Pero las hay
inhibidoras provenientes del cerebro, de la, corteza, y sobre todo del núcleo rojo. Una sección
del neuroeje entre los tubérculos cuadrigéminos anteriores y posteriores suprime esta
influencia inhibidora y produce en el animal un cuadro, denominado rigidez de
descerebración: los cuatro miembros se mantienen rígidamente extendidos, la cabeza y la
columna también extendidas y la cola levantada. El animal es incapaz de moverse y de
cambiar de postura. Si se lo apoya sobre sus cuatro miembros permanecerá erecto durante
largo tiempo, constituyendo esta postura una especie de caricatura de la postura de pie'
normal.
Del juego recíproco de estos dos grupos de influencias depende la regulación del tono
muscular.
Se han descrito numerosos reflejos posturales: algunos, que actúan estando el cuerpo en
reposo (reflejos estáticos), mantienen la postura y la restablecen; otros actúan estando el
cuerpo en movimiento (reflejos estatocinéticos). Todo cambio de postura pondrá en acción a
receptores propioceptivos o exteroceptivos de donde partirán reflejos compensadores, al
mismo tiempo que dejan de excitarse receptores que mantenían la postura anterior.
A) EL LABERINTO. Existen receptores propioceptivos muy importantes que no hemos
mencionado hasta ahora y que se hallan ubicados en el interior del cráneo y forman parte del
oído interno. Es el laberinto, que está constituido por dos partes: una formada por los tres
conductos semicirculares que se disponen en tres planos que están ubicados formando
aproximadamente entre sí ángulos rectos. La otra formada por dos cavidades, el sáculo y el
utrículo. Ambas partes tienen células sensitivas que son estimuladas de diferente manera
según sea la posición de la cabeza. Los cambios de posición del cuerpo, sobre todo aquellos en
que se perturba nuestra posición con respecto al suelo, producen excitaciones del laberinto
donde inician reflejos que tienen por resultado corregir automáticamente el cambio producido.
B) LA CORTEZA CEREBRAL. Los reflejos de tono y los posturales son integrados por
mecanismos nerviosos que tienen su asiento en los ganglios del cerebro, en el cerebelo y en la
corteza cerebral. La extirpación de la corteza cerebral permite la ejecución de reflejos
posturales y reflejos viscerales, pero nunca el conjunto armónico de movimientos que se ve
cuando funcionan normalmente los centros corticales.
DESCRIPCION ANATOMICA. El cerebro del hombre pesa alrededor de 1 200 gramos. Visto
desde arriba se presenta dividido en dos mitades o hemisferios por una gran cisura
longitudinal. Otras cisuras menos pronunciadas penetran más o menos profundamente en la
superficie formando pliegues o circunvoluciones. Visto de perfil se pueden reconocer dos
cisuras profundas: la cisura de Rolando y la cisura de Silvio, que dividen el cerebro en varios
lóbulos: el lóbulo frontal, y el lóbulo parietal, situados respectivamente por delante y por
detrás de la cisura de Rolando; el lóbulo temporal, situado por debajo de la cisura de Silvio, y
detrás de todo el lóbulo occipital.
La sustancia gris de la corteza está formada por células nerviosas de tipo especial según la
zona que se considere. De acuerdo con la citoarquitectura de la corteza se han delimitado
zonas designadas con números.
Cara externa de la corteza cerebral, con las principales áreas citoarquitecturales de la corteza
piramidal (área 4) y de la extrapiramidal
Excitación eléctrica de la corteza. La excitación eléctrica del área 4 produce movimientos
circunscritos en
el lado opuesto del cuerpo. En esta zona nacen las fibras de la motricidad
voluntaria que van a formar el haz piramidal y por eso se denomina sistema piramidal.
También pueden obtenerse movimientos excitando el área 6, llamado premotriz; los
movimientos circunscritos que se obtienen son debidos al paso de estímulos de la zona
excitada al área 4, pues se suprime la respuesta si se extirpa esta zona o se seccionan sus
conexiones nerviosas con el área 6. Pero si el estímulo es más intenso se producirán
movimientos difusos y complejos aun en ausencia del sistema piramidal. El área premotriz es
el componente principal del sistema extrapiramidal, que tiene a su cargo el control de los
movimientos complejos adquiridos por adiestramiento. Este sistema tiene conexiones con el
mesencéfalo y con el cerebelo.
Región prefrontal. (Areas 9, 10, 11, 12 y 13.) El polo frontal adquiere un particular desarrollo
en el hombre. Su excitación no provoca fenómenos motores ni sensoriales. Pero en cambio su
destrucción provoca en el hombre alteraciones importantes de orden intelectual: pérdida de la
capacidad de atención y de retener hechos recientes, facilidad de distracción, incapacidad de
prever. todo lo cual da a estos individuos un aspecto pueril.
Funciones del cerebelo. El cerebelo es una formación nerviosa situada por debajo del cerebro.
Visto desde arriba, después de extirpar la parte posterior del cerebro, se observa que su
superficie presenta circunvoluciones y fisuras menos pronunciadas que las del cerebro.
Consiste como éste en dos hemisferios, conectados por una porción central saliente
denominada vermis.
Cerebelo visto desde arriba después de extirpar la parte posterior del cerebro. Se puede
observar los pedúnculos cerebrales y el vermis.
Es una estación importante en las vías propioceptivas. Sus principales conexiones se
establecen con los núcleos protuberanciales y por su intermedio con la zona premotriz de la
corteza. Interviene en la regulación del tono y de los reflejos posturales sobre los cuales ejerce
una influencia inhibidora, y es un coordinador de los movimientos voluntarios.
El electroencefalograma. La actividad de las neuronas que en número enorme constituyen la
corteza cerebral se manifiesta por latidos o pulsaciones eléctricas rítmicas, debidas a descargas
eléctricas sincrónicas en varias neuronas, que se pueden captar desde la superficie del cráneo y
registrarse previa amplificación.
El trazado resultante se denomina electroencefalograma, y guiándose por la frecuencia y
forma que adopten las ondas eléctricas se pueden localizar tumores cerebrales o diagnosticar
anomalías del funcionamiento cerebral, tales como la epilepsia.
Fisiología de las sensaciones
Los organismos superiores captan del medio ambiente en que viven y de ellos mismos,
diversas impresiones que utilizan para mantener su unidad orgánica y para regular su
conducta. El sistema nervioso es el encargado de este papel informativo. Para ello varias partes
del mismo se especializan en la recepción de determinados estímulos y constituyen los
llamados aparatos sensoriales, que son la vista, el oído, el olfato, el gusto, y las sensibilidades
(superficial y profunda) ya tratadas.
Cada uno de estos aparatos se compone de: a) un receptor periférico con estructura y
características propias; b) un conjunto de fibras nerviosas (vías aferentes) por las cuales van
impulsos de los receptores a los centros; y c) centros donde los impulsos se elaboran y se
asocian (centros superiores) con otros impulsos, resultando la sensación y la respuesta, esta
última por la descarga de impulsos desde los centros a los efectores (músculos o glándulas).
PROPIEDADES GENERALES DE LAS SENSACIONES
Los órganos receptores son de varios tipos, cada uno con estructura y distribución propias y
preferencia para captar pequeñísimas cantidades de una clase de estímulos (irritabilidad
específica). El ojo capta las radiaciones luminosas; el oído las ondas sonoras; la lengua las
sustancias sápidas; la nariz las olorosas; la piel el contacto, la temperatura y el dolor, y las
estructuras profundas, los estímulos de presión, etc.
Estímulos distintos al específico pueden excitar a un receptor, siempre que sean muy intensos,
pero todos suministrarán la misma sensación (ley de Müller). Por ejemplo, si a la retina se le
hace llegar calor, electricidad, o se la toca, dará una sensación luminosa; si lo mismo se repite
con el oído, la sensación percibida será auditiva, y así sucesivamente.
Al estudiar cada sentido en particular se señalarán las características que debe tener el
estímulo para ponerlo en acción, es decir la intensidad mínima y la máxima percibidas, su
duración mínima, la posibilidad de percibir sus distintas variaciones, etc.
Sólo se indicarán aquí algunos principios generales del funcionamiento de los receptores. La
sensación crece al aumentar el estímulo aunque no en igual proporción. Los sentidos localizan
al estímulo, es decir que la sensación se refiere a un punto dado del cuerpo, como cuando se
toca la piel, o fuera del cuerpo, como cuando se señala el lugar de origen de un sonido. Los
sentidos se adaptan fácilmente al estímulo si actúa por mucho tiempo y por ello deja de ser
percibido, como por ejemplo el caso del anillo o el sombrero que por el hábito de llevarlos ya
no se sienten; la adaptación es menos pronunciada al dolor. Además, los sentidos presentan
contrastes, porque disminuye la sensibilidad al estímulo que está actuando y se vuelven más
sensibles al opuesto; así, el ojo que mira con fijeza a un disco rojo sobre fondo blanco se hace
menos sensible al rojo, y si mira en seguida a una superficie blanca verá el color
complementario, es decir un disco verde. Las sensaciones tienen tono afectivo, es decir que
pueden ir acompañadas de un estado de placer o de desagrado. Finalmente los receptores se
fatigan y dejan de responder a un estímulo.
Los receptores se clasifican en dos grupos según su ubicación: exteroceptores, los que se hallan
en la superficie corporal y son capaces de recibir estímulos que llegan de lejos (vista, oído) o de
cerca (gusto, tacto, térmico), e interoceptores, los repartidos en las vísceras o en los huesos,
músculos, etc.
Se examinarán por separado cada uno de los aparatos sensoriales, comprendiendo el estudio
del estímulo, su receptor específico y las vías y centros nerviosos que le pertenecen.
La vista
El sentido de la vista nos permite reconocer los objetos que nos rodean en su forma, tamaño,
color, etc. Sus informaciones contribuyen de manera importante para mantener nuestra
postura corporal y para guiar nuestra marcha. Los ojos son el dispositivo especializado de que
se vale el animal para captar la energía luminosa.
ANATOMIA DEL OJO HUMANO
El ojo tiene la forma de un globo y está situado dentro de la órbita, excavación ósea del cráneo.
Entre el ojo y la órbita se hallan nervios, vasos, grasa, y un conjunto de músculos encargados
de la motilidad ocular.
En la parte anterior del ojo se hallan los párpados, que son repliegues cutáneos móviles; por su
adosamiento impiden la llegada de la luz.
Un corte horizontal del ojo lo muestra constituido por diversas estructuras con funciones
propias.
Corte esquemático horizontal del ojo derecho del hombre, con la indicación de sus diversas
partes.
Rodea al ojo, a modo de cáscara, una envoltura resistente, la esclerótica, con una abertura en
su parte anterior de unos 12 nula de diámetro, en la cual se inserta una formación transparente
combada como un vidrio de reloj, la córnea, por donde penetra la luz.
Detrás de la córnea, pero separado de ella por un espacio lleno de un líquido claro, se halla el
iris, diafragma muscular con abertura central que se agranda o achica permitiendo el mayor o
menor pasaje de luz. Detrás del iris se encuentra el cristalino, que es una lente biconvexa,
transparente, cuya curvatura varía de acuerdo con las necesidades, y modifica así el curso de
los rayos luminosos.
Adosada a la parte interna de la esclerótica se encuentra una capa vascular y pigmentada, la
coroides, y más adentro otra muy delgada, sensible a los rayos luminosos, la retina. Entre la
retina y la cara posterior del cristalino queda una gran cavidad central llena de una sustancia
gelatinosa, transparente, llamada humor vítreo.
LA SENSACION VISUAL (IMPORTANCIA Y CARACTERISTICAS)
Las sensaciones visuales dependen de las propiedades de la radiación luminosa y de las
características del ojo como medio receptor.
A) CARACTERISTICAS DEL ESTIMULO. VISION DE LOS COLORES. Para tener una
sensación visual es necesario que el estímulo luminoso llene ciertas condiciones de intensidad,
duración y longitud de onda.
La intensidad mínima de luz que se puede percibir se llama umbral de intensidad; el umbral
es más bajo cuando se está en la oscuridad. Entre el umbral y la intensidad máxima soportable,
como la del sol, hay una diferencia tan enorme como 1:20.000 millones. El estímulo luminoso
debe durar un tiempo mínimo para poder ser visto; el tiempo es menor cuanto mayor es la
intensidad de la luz.
La luz se propaga por medio de ondas cuya longitud varía para cada color. La longitud de
onda se mide en unidades tan pequeñas como el millonésimo de milímetro o m El ojo sólo
percibe las comprendidas entre 900 y 330 m es decir los colores del espectro solar que van del
rojo al violeta (rojo, anaranjado, amarillo, verde, azul, índigo y violeta). Las radiaciones que
están fuera de estos límites no son captadas por el ojo, aunque pueden ser registradas por el
papel fotográfico y otros dispositivos. Existe un umbral particular para cada color, siendo el
del amarillo verde el más bajo de todos. El ojo distingue unos 180 tonos, pero puede no
especificar si son puros o mezclas.
Algunas personas tienen ceguera para uno o más colores. En homenaje a Dalton (1774), que la
padecía y fue el primero en señalarla, se la denomina daltonismo. Por lo común esta ceguera
es parcial y afecta a los colores verde y rojo, que no pueden ser distinguidos; este defecto es un
inconveniente grave en ciertos trabajos (maquinistas, marinos). Existen medios muy sencillos
para descubrirla y entre ellos las tablas de Ishijara, en las que el sujeto mira un rompecabezas
formado por círculos de varios colores y ve un número, que es distinto en los de visión normal
v en los daltónicos. Por procedimientos como éste o análogos, se observa que este trastorno
existe en el 2% de las mujeres y en el 9% de los varones sanos, y que la mayor parte ignora que
lo posee.
CARACTERISTICAS DE LA PERCEPCION POR EL OJO.
Hay varias particularidades en la
visión normal, entre ellas algunas que a veces inducen a error.
El campo visual es la zona que un ojo inmóvil puede ver. Su conocimiento es de interés, pues
para el ojo normal tiene una extensión dada cuyas reducciones son índice de ciertas
enfermedades.
Se llama agudeza visual la capacidad de distinguir 2 puntos separados pero próximos entre sí.
De ella depende la posibilidad de ver detalles. Al observar las estrellas algunas personas
distinguen dos, donde para otras hay sólo una. Por regla general dos objetos deben hallarse
separados por un ángulo de I minuto para poder distinguir uno del otro; el ángulo está
formado por dos líneas rectas que van del ojo del observador a cada uno de los puntos. La
agudeza visual se mide, en la práctica, colocando al sujeto frente a un cartel con letras de
diversos tamaños y pidiéndole que lea las más pequeñas que pueda observar.
En el ojo hay persistencia de las imágenes hasta más de 20 a 30 milisegundos después de haber
actuado el estímulo. Esto tiene una feliz aplicación en el cine, donde proyecciones separadas
dan por esa persistencia la sensación de continuidad y movimiento. Por igual razón un punto
que se mueva rápidamente, como una estrella fugaz, parece ser una línea.
Ilusiones ópticas. Se denominan así a las impresiones visuales que infieren un juicio falso
sobre la forma, tamaño, etc., de los objetos observados.
Ilusión óptica. En este grabado todas las líneas son círculos de diámetro cada vez mayor; el
contraste entre blanco, negro y gris, produce la ilusión de que se trata de espirales. Podemos
comprobar que somos víctimas de una ilusión óptica, siguiendo con un lápiz el contorno de las
supuestas espirales.
MARCHA DE LOS RAYOS LUMINOSOS EN EL OJO
El ojo puede ser comparado con una cámara fotográfica; como ella recibe la luz y por un
sistema óptico especial forma las imágenes en la retina. Un grupo de anexos al sistema óptico
facilita o, mejor dicho, hace posible su correcto funcionamiento.
ANEXOS DEL SISTEMA OPTICO DEL OJO. a) Movimientos oculares. El globo ocular ejecuta
movimientos de rotación dentro de la cavidad orbitaria en la que se aloja, gracias a la acción
combinada de 6 músculos gobernados por la voluntad. Estos músculos se insertan en la pared
de la órbita y en el ojo, y sus movimientos están coordinados entre sí y con los músculos del
otro ojo, por el sistema nervioso.
La visión binocular, o sea el hecho de que observando un objeto con los dos ojos se vea simple
y no doble, se debe al juego combinado de la musculatura de ambos ojos, que los desvían
hasta que las imágenes se forman en puntos simétricos de ambas retinas. Por eso, desviando el
eje de uno de los ojos por presión ejercida con el dedo sobre el globo ocular, la imagen del
objeto aparecerá doble. La visión binocular es importante ya que por ella se aprecia el relieve y
la profundidad de los objetos.
b) Protección y nutrición del ojo. Por delante de los ojos se hallan los párpados, formados por
una lámina cartilaginosa recubierta adelante por la piel y atrás por la conjuntiva. En su espesor
y rodeándolos hay un músculo circular que al contraerse cierra los párpados. En el párpado
superior se inserta un músculo que lo levanta, el elevador. El cierre de los párpados se puede
realizar por acción de la voluntad o en forma refleja ante estímulos diversos como la luz
intensa, el contacto de cuerpos extraños con el ojo, o durante el sueño. El parpadeo habitual se
realiza para humedecer con lágrimas a la córnea, y su estímulo es la desecación de la superficie
de ésta.
Las lágrimas son el producto de secreción de las glándulas lagrimales situadas en el espesor
del párpado superior. Son un líquido acuoso que contiene sal, bicarbonato y algo de albúmina.
Su papel es humedecer la parte expuesta del ojo, córnea y conjuntiva, y barrer los gérmenes
que se depositen en ellas. Dos pequeños orificios situados cerca de la comisura interna de los
párpados les dan salida hacia un conducto único que desemboca en la nariz.
Las arterias que nutren el globo ocular son: la central de la retina, que llega junto con el nervio
óptico y es visible en el ser vivo iluminando el interior del ojo con medios adecuados, y,
además, varias otras derivadas de la oftálmica, que se reparten entre la esclerótica y la retina.
EL SISTEMA OPTICO DEL OJO. Se analizarán los diversos medios que atraviesa la luz que
viene del exterior para alcanzar la retina y formar allí la imagen luminosa.
a) Córnea. Está engarzada en el orificio circular y anterior de la esclerótica, y tiene la forma de
un vidrio de reloj curvo con sus dos caras paralelas. Es muy transparente a los rayos
luminosos. Se la ve a simple vista y su curvatura se aprecia bien mirándola de costado.
La deformación de alguno de sus meridianos deforma la imagen retiniana al no coincidir el
foco de ese plano con el de los otros. A este defecto se le llama astigmatismo y se lo corrige con
lentes.
b) Humor acuoso. La pequeña cavidad situada entre la cara posterior de la córnea y la anterior
del cristalino, y dentro de la cual emerge el iris, como un diafragma, se halla ocupada por el
humor acuoso. Este es un líquido transparente, con composición análoga a la del plasma
sanguíneo luego de filtrado a través de poros muy finos; esto se explica, pues tiene origen en la
sangre por filtración capilar. Se halla a una presión de 25 a 30 mm de mercurio, la que
aumenta en el glaucoma, y produce entonces dolores intolerables.
c) Iris. Los rayos luminosos pasan al interior del ojo a través de la pupila, orificio circular que
está en el centro del iris. Este es una membrana de distintos colores, según las personas. azul,
castaño, etc., situada delante del cristalino y que funciona como un diafragma fotográfico.
Posee en su interior dos músculos: uno circular, cuya contracción disminuye el diámetro de la
pupila reduciendo así la cantidad de luz que llega a la retina; y otro dispuesto en forma
radiada, cuya contracción aumenta el diámetro de la pupila permitiendo la llegada de mayor
cantidad de luz a la retina. En sus posiciones extremas la pupila varía de 1 a 8 mm de
diámetro.
Los movimientos pupilares escapan a la voluntad, pues son reflejos gobernados por el sistema
nervioso. Por ese mecanismo la pupila se acomoda a la luz, agrandándose en la oscuridad y
achicándose por la iluminación, y se acomoda a la distancia, dilatándose al mirar los objetos
lejanos y reduciéndose ante los cercanos. El diámetro pupilar también es modificado por la
acción de algunas drogas que actúan sobre los nervios reguladores de ese diámetro.
ACCION DE ALGUNOS AGENTES SOBRE EL DIAMETRO DE LA PUPILA
(*) La belladona fue usada primeramente por las mujeres venecianas para tener ojos con un
brillo particular que realzara su belleza. Su principio activo es la atropina, muy usada por los
oculistas para poder observar con comodidad el fondo del ojo.
d) Cristalino. El cristalino es una lente biconvexa fijada por su borde cerca del músculo ciliar
que la rodea y bañada en su cara anterior por el humor acuoso y en la posterior por el humor
vítreo. Es el pasaje obligado de todos los rayos luminosos, a los que concentra en la retina.
Su propiedad fundamental es la de deformarse, haciendo sus caras más o menos curvas, según
la distancia a que mire el sujeto. Esta acomodación a la distancia se logra por el grado variable
de tracción que ejerce el músculo ciliar sobre el borde de inserción del cristalino. Esta
deformación es sobre todo de su cara anterior y puede apreciarse a simple vista en las
llamadas imágenes de Purkinje, colocando al sujeto en una habitación a oscuras y acercando
una vela a su ojo.
Imágenes de Purkinje en el ojo que mira a lo lejos (A), y en el ojo que mira de cerca (B). El
sujeto se halla en la oscuridad y se le acerca tina vela cuyas imágenes corresponden: a y a' en la
córnea, b y b' en la cara anterior del cristalino, c y e' en la cara posterior del mismo.
Toda vez que se mira a un objeto cercano al ojo, la cara anterior del cristalino se encorva
exageradamente y por el contrario, se endereza, si el objeto se aleja. La finalidad de estos
movimientos es que la imagen se haga siempre nítida en la retina. Pero hay dos límites
extremos dentro de los cuales se manifiesta la acomodación a la distancia del cristalino, los que
coinciden con el punto próximo y el punto remoto.
Punto próximo es la distancia mínima a que podemos distinguir claramente un objeto.
Aumenta con la edad; a los 20 años es de unos 10 cm, a los 40 años de unos 22 cm, y a los 60
años de un metro. Se llama presbicia, a este exagerado alejamiento observado luego de los 40
años. La distancia mínima a que puede observarse un objeto sin la acomodación del cristalino
se llama punto remoto; más allá del cual los rayos luminosos son prácticamente paralelos. Es
de unos 6 metros. Vale decir entonces, que el poder de acomodación del cristalino se ejerce
cuando se miran objetos comprendidos entre distancias de 10 cm y 6 metros.
Cuando el cristalino es incapaz de acomodar el ojo a la distancia, se corrige el defecto
utilizando lentes de tipo adecuado al defecto. Esto sucede en la presbicia; en la miopía el punto
próximo está muy cercano y el sujeto ve bien de cerca y mal de lejos por formarse la imagen
delante de la retina; en la hipermetropía, el punto próximo se halla alejado y el sujeto necesita
acomodar el ojo para ver a lo lejos, puesto que la imagen se forma detrás de la retina.
Marcha de los rayos luminosos y formación de la imagen en la retina, en ojos normales y con
trastornos de convergencia de los rayos luminosos. Corrección por lentes.
En algunas circunstancias, el cristalino se opacifica y lleva a la ceguera al no permitir el pasaje
de los rayos luminosos. La catarata, como se denomina esta afección, se observa en la vejez, la
diabetes, etc. Una operación que elimine la parte opaca puede restituir la visión.
e) Retina. La retina es la delgada membrana que recubre por dentro al globo ocular. Por su
cara externa apoya sobre la coroides, membrana muy vascular y por la interna la baña el
cuerpo vítreo, medio transparente que le lleva los rayos luminosos. Al nivel del eje
antero
posterior del ojo presenta una pequeña depresión, la fovea centralis, de importancia capital ya
que es el lugar que proporciona la visión más perfecta. Unos milímetros más adentro de la
fovea
se halla una protuberancia llamada papila, que corresponde a la entrada del nervio
óptico en el ojo. Por la papila penetra, junto con el nervio óptico, la arteria central de la retina
cuya misión es irrigar a esta membrana. Iluminando el fondo del ojo con el oftalmoscopio es
posible observar la papila, la arteria central de la retina y la fovea.
Estudiada en un corte microscópico la retina presenta una estructura compleja, formada por
enorme cantidad de células y fibras nerviosas. En su parte más extensa se destacan dos tipos
particulares de células sensibles a la luz, los conos y los bastoncillos, cuyo nombre da una idea
de su forma, y que se apoyan por fuera en una capa pigmentaria.
La función especializada de la retina es transformar la energía luminosa en energía nerviosa,
que por los nervios envía al cerebro. Esta función depende de la presencia de los conos y
bastoncillos, ya que la papila que no contiene a ninguno de ellos, es ciega.
Punto ciego de la retina. Cerrar el ojo izquierdo y mirar fijamente la cruz, acercando y alejando
la figura; se verán tanto la cruz como el círculo blanco, hasta que en determinado momento
desaparecerá este último, cuando se forme su imagen en la retina (a unos 20 cm.).
Los conos son los que permiten la nitidez de la percepción y la visión de los distintos colores.
La visión a nivel de la fovea llena estas condiciones y en ella
sólo hay conos. Los conos
necesitan para funcionar una buena iluminación y por eso son apropiados para la visión
diurna. Faltan en las aves y mamíferos nocturnos (murciélago, erizo, topo) y se hallan muy
desarrollados en las aves diurnas que buscan insectos de colores vivos.
Los bastoncillos están adaptados a la visión crepuscular. Perciben intensidades débiles de luz
pero dan imágenes borrosas como la de bultos que se desplazan y no permiten conocer el
color. Es el tipo de la visión nocturna. La parte periférica de la retina, alejada de la fovea y rica
en bastoncillos, es la encargada de este tipo de visión. Es interesante señalar que alrededor de
la extremidad alejada de los bastones se halla un pigmento rosado, la rodopsina, que se
descolora por acción de la luz y estimula al bastoncito; y en la oscuridad recupera su color
siempre que la persona tenga suficiente vitamina A. La falta de vitamina A en la alimentación
de los aviadores, por ejemplo, les crea serias dificultades y a veces es causa de accidentes al
levantar vuelo o aterrizar con luz escasa.
LAS VIAS Y LOS CENTROS NERVIOSOS VISUALES
Al nervio óptico llegan todas las fibras nerviosas especializadas que vienen de la retina. Estas
fibras traen todas las impresiones recogidas por los conos y bastoncillos, salen del ojo por la
papila y se dirigen al encuentro del nervio óptico del lado opuesto, con el que se fusionan en
el quiasma óptico, situado en la base del cerebro. De aquí parten las cintillas ópticas, una por
cada lado, que después de hacer escala en el cuerpo geniculado externo, penetran en el
cerebro y se abren en abanico y reparten en la corteza del lóbulo posterior del cerebro (ambas
márgenes de la cisura calcarina del lóbulo occipital).
Esquema que representa el camino de las vías ópticas desde el ojo hasta la corteza cerebral
occipital, y su punto de entrecruzamiento al nivel del quiasma.
La corteza cerebral occipital es la etapa final de los estímulos retinianos y constituye su centro
visual. La corteza del lado izquierdo del cerebro recibe las impresiones que parten de la mitad
izquierda de los dos ojos, y la corteza del lado derecho las impresiones que parten de la mitad
derecha. La razón está en que al nivel del quiasma se entrecruzan las fibras más internas de los
nervios ópticos y pasan al lado opuesto. Teniendo presente que un cuerpo situado a la derecha
del observador se proyecta en la mitad izquierda de las dos retinas, cuyas fibras se encuentran
luego en el quiasma y siguen juntas, se explica, 1) por qué el notable entrecruzamiento de las
vías ópticas, y 2) por qué el cerebro izquierdo es el que ve los objetos situados a la derecha y
viceversa.
El curso tan particular de las vías ópticas explica también por qué un corte del nervio óptico da
ceguera sólo en ese ojo; por qué si el corte es antero posterior y divide el quiasma, la ceguera
ataca la mitad nasal de las retinas; y por qué el corte de la cintilla óptica deja ciega la mitad de
ese lado de ambos ojos.
La audición
El oído reproduce con fidelidad los sonidos que le llegan y así aporta al organismo
valiosísimas in formaciones del medio ambiente. El habla, que permite las relaciones con los
semejantes, se aprende por su intermedio. Los sonidos y los ruidos despiertan sensaciones
agradables o desagradables, por lo que se dice que este sentido tiene tono afectivo.
La fisiología del oído se halla en plena revolución por la aplicación a su estudio de las nuevas
técnicas electromecánicas de producción, amplificación y re. producción de los sonidos,
usadas en la radio, el cine, la fonografía, etc.
LA SENSACION AUDITIVA
Se considerará separadamente teniendo en cuenta las variaciones que dependen del estímulo
en sí y las que son propias del órgano receptor.
A) CARACTERISTICAS DEL ESTIMULO. El sonido consiste en vibraciones que se propagan
por los medios sólidos, líquidos o gaseosos. Sus propiedades son: la intensidad (fuerte o débil),
el tono o
altura (agudo o grave), y el timbre, que permite reconocer en una misma nota al
instrumento que la da.
Dentro de ciertos límites el oído humano reconoce las tres propiedades. El tono de un sonido
depende de su número de vibraciones por segundo y el oído capta frecuencias comprendidas
entre 16 y 20.000 vibraciones. La voz humana tiene entre 300 y 3.000, y en música se suelen
usar notas que oscilan entre 25 y 4.800 vibraciones por segundo. Cada especie animal oye
determinadas frecuencias de sonidos; así, la rata y el perro, perciben sonidos agudísimos,
inaudibles para nosotros. El murciélago guía su vuelo nocturno entre obstáculos por gritos de
esa índole, cuyo eco recibe su oído, tal como funciona con ondas de radio un aparato de radar.
La intensidad mínima de un sonido capaz de ser oído varía con el tono. Así, el summum de
audibilidad se halla en las 2 700 vibraciones por segundo y más arriba o más abajo, la
intensidad debe aumentar para ser perceptible. Este mínimo aumenta pasando los 30 años de
edad, en particular para los tonos agudos. La intensidad máxima audible es cercana a la del
trueno y ocasiona molestias al que escucha.
B) CARACTERISTICAS DE LA PERCEPCION
POR EL OIDO. Se llama poder analítico
del
oído a su facultad de distinguir los
componentes de un sonido complejo. En los músicos se
halla muy desarrollado y entre otras cosas
les permite individualizar
a varias notas que
suenan
juntas, lo que exigiría
complicadas operaciones
con otros procedimientos.
El oído
localiza el origen de la fuente sonora. Este reconocimiento lo hace ayudado por movimientos
de la cabeza y por las diferencias de percepción del mismo sonido en
ambos oídos. Ciertos
dispositivos para localizar aviones en la noche imitan este mecanismo. Junto con la vista, es el
oído
un sentido proyectivo, ya
que localiza fuera del
cuerpo al fenómeno originario de la
sensación.
MECANICA DE LA AUDICION
Se acostumbra a subdividir el estudio de la función auditiva en tres partes, oído externo,
medio e interno, lo que se halla justificado por la anatomía, la fisiología y la patología.
El oído externo comprende las orejas, que reciben las vibraciones sonoras, y un tubo de 2,5 cm
de largo por 0,7 cm de ancho, tortuoso, que orienta esas vibraciones hacia el tímpano. En la
entrada de este conducto unos pelos y un poco de cerumen lo protegen.
Corte vérticotransversal del aparato auditivo, en el cual se destacan con claridad el oído
externo, medio e interno, y demás partes.
El oído medio y el interno, que continúan al anterior, se hallan situados en pleno hueso
peñasco (dependencia del hueso temporal), en la base del cráneo. Se examinarán por separado.
OIDO MEDIO. Comprende la caja del tímpano, cavidad de 1 a 2 cm situada en el peñasco y
sus anexos. Por fuera se halla separada del conducto auditivo descrito, por la membrana del
tímpano, que es elíptica, de 10 X 9 mm, muy inclinada y combada hacia adentro. Por dentro
tiene una pared ósea con dos ventanitas, la oval y la redonda, que la comunican con el oído
interno. Por abajo y por delante se prolonga en un tubo que la comunica con la parte posterior
de la nariz, la trompa de Eustaquio. En su parte posterior desembocan por unos orificios las
cavidades de la apófisis mastoides. Finalmente, en su interior hay una cadena formada por tres
minúsculos huesecillos, que pesan en conjunto unos 55 mg, y se extienden como puente desde
la pared externa a la interna, es decir, desde la membrana del tímpano a la ventana oval.
La fisiología del oído medio es la siguiente: las vibraciones sonoras que llegan por el conducto
auditivo ponen en vibración a la membrana del tímpano, que es muy sensible y reproduce el
sonido con fidelidad. Los extremos de la cadena de huesecillos ponen en comunicación la
amplia membrana del tímpano (90 mm2 de superficie) con la pequeña ventana oval (3 mm2 de
extensión) transmitiendo, de este modo, concentradas, las vibraciones de la primera a la
segunda.
La destrucción de los huesecillos produce las llamadas sorderas de transmisión, en las cuales
la audición de sonidos que llegan por el aire está imposibilitada, pero se pueden oír los
sonidos que ponen en vibración a los huesos del cráneo y que por su intermedio llegan al oído
interno, como por ejemplo un diapasón apoyado en la cabeza.
La trompa de Eustaquio, que constituye una comunicación entre la caja del tímpano y el
exterior por vía de la nariz, permite que la membrana del tímpano tenga sus dos caras bajo la
presión atmosférica y vibre en condiciones óptimas. Basta la obstrucción de la trompa por
efecto de un resfriado, para que se produzca un desequilibrio y una ligera sordera.
OIDO INTERNO. El oído interno, junto con la porción correspondiente del sistema nervioso,
son las partes esenciales del aparato auditivo. Su destrucción produce sorderas incurables
llamadas de recepción.
A la izquierda y abajo, el caracol con sus espiras; en punteado el lugar del corte transversal,
que aparece muy ampliado a la derecha. En dicho corte se ven las dos rampas y el conducto
corlear, así como las demás partes.
El caracol o coclea es el receptáculo anatómico del oído interno. Está formado por un tubo óseo
situado en pleno peñasco, enrollado 21/2 veces como la envoltura de aquel animal. La base del
caracol mide 9 mm, y comunica con la ventana de la pared del oído medio; su altura es de sólo
5 mm. Su cavidad se halla recorrida a lo largo por dos membranas que la dividen en tres
conductos independientes y superpuestos, llenos de linfa. La membrana del conducto
intermedio, llamado conducto
coclear, que mira hacia la base del caracol, se denomina
membrana basilar y sobre ella apoya el órgano de Corti, formado por células de un tipo y
disposición articulares que reciben una rica inervación. La membrana basilar y el órgano de
Corti son los dispositivos especializados que captan las vibraciones debidas al sonido.
El funcionamiento del oído interno es el siguiente: Las vibraciones de los huesecillos
trasmitidas a la ventana oval se comunican a la linfa, que llena el caracol, produciendo
variaciones de presión en este líquido, variaciones que son captadas por la membrana basilar y
el órgano de Corti. Pero cada región de esta membrana se halla especializada en la recepción
de vibraciones de determinada frecuencia; las cercanas a la base del caracol son sensibles a los
tonos agudos y su lesión perturba la audición de éstos, como sucede en los viejos, y las
cercanas al vértice son sensibles a los tonos graves. Es decir que hay una localización del oído
interno para la recepción de cada tono.
En el año 1930, Weber y Bray llamaron la atención sobre el curioso hecho de que cada vez que
llega una vibración al oído interno, éste produce corrientes eléctricas que se pueden captar
como las ondas de la radio y retransformar en sonido. Por eso, para precisar su origen, se las
llama corrientes microfónicas cocleares. El experimento de estos investigadores ingleses es
sorprendente. Conectaron dos hilos de un aparato receptor de radio con el oído interno de un
gato y los pusieron en comunicación con un altavoz situado en una habitación alejada; bastaba
hablar quedamente en la oreja del gato, para que el altavoz repitiera las palabras.
VIAS Y CENTROS NERVIOSOS
El nervio coclear, rama del auditivo, nace en el caracol alrededor de las células del órgano de
Corti y se dirige al bulbo raquídeo en el que penetra. De allí asciende, luego de cruzar la línea
media, para llegar a su punto terminal en la corteza del cerebro, en su lóbulo temporal, es
decir en la parte lateral. Este largo recorrido de las fibras se hace en cuatro etapas sucesivas,
cada una a cargo de una neurona. El estímulo nervioso sufre un retardo acentuado al pasar de
una fibra a la siguiente.
La corteza del lóbulo temporal de cada lado constituye el centro de la audición, que recibe
fibras nerviosas del oído del lado opuesto y del mismo lado. Por eso son reemplazables el uno
por el otro. Los tumores destructores de un solo centro o su extirpación, dificultan la audición.
Su excitación por medio de toques o corrientes eléctricas hace percibir ruidos variados.
Los sentidos químicos: el olfato y el gusto
Estos dos sentidos tienen receptores especializados que son excitados por agentes que para
actuar deben disolverse en la saliva o secreción nasal. Las sensaciones que ambos suministran
suelen ir asociadas y basta un resfriado para notar el cambio de sabor de algunos alimentos.
EL OLFATO
El sentido del olfato tiene tono afectivo, pues los olores que percibimos nos dan una sensación
de placer o desagrado. Cumple un papel iniciador y estimulante de la digestión, pues el olor
de los alimentos inicia y aumenta la secreción de saliva y jugo gástrico. En los animales de fino
olfato es un guía valioso para su defensa, la búsqueda de la presa, y también para localizar al
otro sexo.
Sus elementos receptores se hallan situados en la parte alta de las fosas nasales, en una
superficie amarillenta de unos 250 mm2. Allí, entre células de sostén de la mucosa pituitaria,
se hallan células olfatorias, que perforan la superficie en la que dejan emerger un pincelillo de
6 a 8 pestañas.
Pared externa de las fosas nasales, con las ramificaciones del nervio olfatorio. que se extienden
por el cornete superior y parte del medio.
La sensibilidad del olfato varía notablemente con los olores, las personas y la especie animal.
El estimulante más poderoso para el hombre es el almizcle artificial, el que se huele aun en
concentración de 0,04 de millonésima de gramo por litro; el almizcle natural necesita dosis mil
veces mayores. La medición se hace por dispositivos llamados olfatómetros. Este sentido se
adapta fácilmente a los olores, los que al cabo de un tiempo relativamente corto se perciben
menos.
La sensación olfativa se origina porque las sustancias olorosas desprenden partículas que
llegan a la pituitaria, donde se disuelven en el mucus y así excitan a las células olfatorias. El
estímulo nervioso nacido en estas células entra al cráneo por el filete que envían al bulbo
olfatorio, pasa luego a la cintilla olfatoria y alcanza al cerebro terminando en la circunvolución
del hipocampo (muy desarrollada en los peces) situada en la parte inferior del mismo.
EL GUSTO
Este sentido es un seleccionador precioso de la calidad de los alimentos, y evita en gran parte
la ingestión de sustancias nocivas. junto con el olfato actúa en la digestión, iniciando la
secreción de saliva y de jugo gástrico. (Es bien conocido lo que su educación consigue en el
hombre, pues los grandes gourmets son personas de edad.)
Los receptores del gusto se hallan repartidos por el dorso de la lengua, pared posterior de la
faringe, el velo del paladar y su pilar anterior, pero no en la campanilla. Observados al
microscopio se ven corpúsculos en forma de barril, rodeados de fibras nerviosas y con dos
tipos de células en su interior, unas de sostén y las otras gustativas, con un pelo libre dentro
del corpúsculo.
La capacidad gustativa varía mucho de una persona a otra. Así mientras unos perciben una
solución de azúcar de caña al 0.41%, otros ni siquiera reconocen la del 10%. Hay personas
incapaces de diferenciar la acidez del vinagre, del amargo de la quinina.
Fuera del factor personal señalado se observan fácilmente los contrastes en este sentido. El
agua destilada, que es insípida, parece dulce luego de probar una solución ácida; el azúcar se
percibe en menor cantidad después de tomar algo amargo.
Las sustancias, según la sensación gustativa que proporcionan, se clasifican en cuatro grandes
grupos, sin considerar las sustancias insípidas:
A) SALADAS. Su tipo es la sal de cocina, cuya concentración mínima percibida es de 0,01%.
Las ratas la perciben tanto más fácilmente cuanto menos sal tienen en su sangre. La punta de
la lengua es la parte de la boca especializada para percibir este sabor.
B) ACIDAS. Se perciben con este gusto todos los ácidos siempre que no quemen. En las partes
laterales de la lengua está localizado este tipo de sensación.
C) DULCES. Así saben todos los azúcares, alcoholes y sus derivados, y otras
sustancias como el
cloroformo, etc. Se las percibe con la punta de la lengua.
D) AMARGAS. Son los alcaloides (morfina, estricnina, cocaína, quinina, etc.), los glucósidos de
la digital, almendras amargas, etc., y otras como las sales biliares. La inyección en la sangre de
decolina da sabor amargo en la boca. Esta sensación se localiza en la base de la lengua.
Las partes rayadas indican las regiones de la lengua donde se localizan los distintos sabores.
Las sustancias gustativas para ejercer su acción deben venir disueltas o disolverse en la saliva
y ponerse en contacto con los corpúsculos del gusto antes señalados. Cada tipo de sustancias
estimula solamente a su correspondiente categoría de receptor, de los que hay cuatro. El
estímulo nervioso así nacido sigue por los nervios lingual, glosofaríngeo y vago, llega al bulbo
raquídeo, pasa al tálamo y termina en la corteza parietal, del cerebro del lado opuesto.
Sensibilidad superficial y profunda
Repartidos en la piel y diversas estructuras profundas (huesos, músculos, vísceras, etc.) existen
varios receptores sensoriales, que difieren por su estructura, el estímulo que los pone en acción
y las vías nerviosas que les corresponden. Se estudian por separado los ubicados en la piel y
los ubicados en la profundidad del cuerpo.
SENSIBILIDAD SUPERFICIAL
En la piel se observan puntos sensibles al tacto, otros a los cambios de temperatura y otros al
dolor, repartidos en forma desigual pero no superpuestos. A cada uno de estos puntos
cutáneos corresponde un receptor especializado.
Los tres tipos de receptores presentan
dos particularidades comunes: localizan can bastante
exactitud el punto de la piel sobre el cual actúa el estímulo y, si éste no es muy poderoso, se
adaptan a él y deja de sentirse, con excepción de los estímulos dolorosos. La adaptación
explica por qué no se sienten los vestidos.
SENTIDO DEL TACTO. Su estimulo es la deformación de la piel en un punto o zona; por eso
al sumergir un dedo en mercurio sólo se siente el borde que forma éste alrededor del dedo.
Sus receptores están formados por una fibra nerviosa situada en el espesor de la piel, que se
divide y ensancha en las terminaciones (discos de Merkel), o se enrosca en una espiral rodeada
por tejido conectivo formando un corpúsculo (Meissner), o abraza la base de un pelo en forma
de cesta.
Receptores cutáneos. Arriba y a la izquierda: corpúsculo de Krause; arriba y a la derecha: disco
de Merkel; abajo y a la izquierda: terminaciones nerviosas libres en la córnea; abajo y a la
derecha: corpúsculos de Pacini.
La mayor concentración de estos receptores se halla en los sitios donde se comprueba máxima
sensibilidad: dedos, punta de la lengua, labios.
Sensibilidad de la piel al tacto medida por: A) el peso mínimo en gramos percibido; y B) la
distancia mínima en mm para percibir como separados dos pinchazos simultáneos.
SENTIDO TERMICO. En la piel hay unos puntos sensibles al calor y otros al frío. Los
receptores del frío son unos corpúsculos redondeados (Krause); los correspondientes al calor
no se conocen bien. Hay 8 puntos sensibles al frío por cada uno sensible al calor.
La sensibilidad térmica varía de una zona a otra del cuerpo, siendo las partes cubiertas las más
sensibles (pecho, abdomen); la falta de circulación también la aumenta. El mentol estimula
específicamente los corpúsculos del frío y da esta sensación.
SENTIDO DEL DOLOR. Se caracteriza
porque va acompañado por una sensación de
desagrado, es decir, tiene tono afectivo elevado. Cuando esta sensación es muy fuerte el sujeto
trata de alejarse del estímulo doloroso.
El estímulo puede ser muy variado, electricidad, temperatura, agentes mecánicos, químicos,
etc., con tal que lesione los tejidos. El dolor es, pues, un índice de lesión tisular.
Sus receptores se hallan no sólo en la piel sino en muchos otros órganos y tejidos como se verá
luego. Estos receptores son simples terminaciones nerviosas, de las cuales hay gran
abundancia en las zonas sensibles; donde faltan no hay dolor.
La sensación dolorosa que tiene lugar en la piel o mucosas es localizada; la profunda es difusa;
a veces se localiza en un punto alejado del lugar de origen. Es de señalar el hecho curioso de
que hay sujetos con ausencia de sensibilidad al dolor (analgesia congénita).
El mecanismo por el cual se produce el dolor es la compresión de las extremidades nerviosas
libres o su ataque químico por sustancias liberadas por las células al ser lesionadas.
SENSIBILIDAD PROFUNDA
Comprende numerosas y variadas sensaciones que nacen en la intimidad de los órganos y
tejidos del cuerpo, y son de mucha utilidad para mantener la unidad funcional y la relación
entre las partes. Se consideran por separado la sensibilidad de las vísceras y la propioceptiva.
SENSIBILIDAD VISCERAL. Los
nervios
viscerales captan continuamente por sus
terminaciones, impresiones que transportan a los centros nerviosos, pero que pocas veces
entran en la esfera de la conciencia. Estas impresiones difieren según el órgano de que se trate.
El corazón y las grandes arterias se informan de los cambios de la presión arterial, del oxígeno
y del anhídrido carbónico de la sangre, y con esa base regulan los movimientos respiratorios y
la misma presión arterial. De la superficie pulmonar y pleural, nacen estímulos que por medio
del nervio vago gobiernan la respiración. De la tráquea, la laringe y la nariz, parten estímulos
que provocan la tos o el estornudo que sirven para limpiar las vías respiratorias. En la vejiga
nacen las sensaciones que provocan la micción.
Aparte de las diversas sensaciones enumeradas también hay sensación de dolor en algunas
vísceras y en otras no. El corazón, que no tiene sensibilidad táctil, la tiene dolorosa, de lo que
es prueba el intenso dolor de la angina de pecho. La pleura que recubre la pared costal y la
diafragmática, duele intensamente y en cambio la que tapiza al pulmón es indolora. Tienen
sensibilidad dolorosa el peritoneo, las vías biliares y las renales, los dientes, etc., como bien lo
saben los pacientes con peritonitis, cólicos, etc.; no la tienen el hígado, el bazo, etc. Un hecho
bien conocido pero de mecanismo todavía desconocido, es que el dolor puede manifestarse
lejos de la víscera enferma (dolor referido).
SENSIBILIDAD PROPIOCEPTIVA. Comprende también un complejo grupo de sensaciones
originadas en zonas profundas del cuerpo, que permiten apreciar y mantienen la postura de
cada una de las partes del organismo, el equilibrio corporal y los movimientos.
El estímulo es una fuerza que por presión o tracción deforma una parte del cuerpo.
Los receptores son varios. Unos se hallan alrededor de las fibras de todos los tendones y
músculos e informan sobre el grado de estiramiento que éstos sufren. Otros, formados por
unos grandes corpúsculos ovales (Pacini) situados en la palma de la mano y el pie, en la
superficie de las articulaciones y los huesos, informan de las presiones sufridas en esos
lugares. Finalmente los del laberinto, situados al lado del oído interno, reciben las variaciones
de presión de la linfa allí contenida, según la posición de la cabeza, y sus informaciones son
utilísimas para la conservación del equilibrio.
VIAS Y CENTROS NERVIOSOS DE LA SENSIBILIDAD SUPERFICIAL Y PROFUNDA
Las fibras nerviosas que corresponden a los distintos y numerosos receptores cutáneos y
profundos, confluyen con las homólogas formando los nervios y se dirigen a la medula espinal
para ascender y, luego de algunas escalas, finalizar en la corteza cerebral. Pero este largo
trayecto ofrece algunas particularidades para cada tipo de sensibilidad.
Las fibras sensitivas de los nervios entran en la medula espinal por las raíces posteriores y
toman ubicación determinada para cada tipo de sensibilidad.
Las fibras sensitivas de los nervios entran en la medula espinal por las raíces posteriores y
toman ubicación determinada para cada tipo de sensibilidad. Las correspondientes a la
sensibilidad profunda se reparten en dos grupos: uno que asciende por los cordones
posteriores de la medula del mismo lado por el que entra, y pasa al opuesto antes de llegar al
tálamo; el segundo asciende por los cordones laterales de ambos lados y se detiene, por
último, en el cerebelo.
Las correspondientes a la sensibilidad profunda se reparten en dos grupos: uno que asciende
por los cordones posteriores de la medula del mismo lado por el que entra, y pasa al opuesto
poco antes de llegar al tálamo, en el que hace escala; el segundo grupo asciende por los cordones
laterales de ambos lados y se detiene en el cerebelo.
Las vías nerviosas de la sensibilidad superficial (táctil, térmica y dolorosa) entran a la medula,
pasan al lado opuesto y ascienden por los cordones antero laterales hasta el tálamo, en el que
se detienen; en la medula las tres sensibilidades, táctil, térmica y dolorosa, se disponen según
cierto plan. En definitiva, todas las vías sensitivas del organismo van a parar al tálamo del
lado opuesto al que provienen, con excepción de un grupo perteneciente a la sensibilidad
profunda que no se cruza y se dirige, pues, directamente al cerebelo.
La distribución tan particular de las vías sensitivas en su trayecto medular explica los síntomas
de algunas enfermedades. En la ataxia locomotriz se lesionan los cordones posteriores y estos
pacientes pierden la sensibilidad profunda y tienen grandes dificultades para mantenerse de
pie o caminar, sobre todo si cierran sus ojos y pierden las sensaciones visuales. En la
siringomielia están destruidas las fibras de la sensibilidad superficial a poco de su entrada, en
su trayecto hacia adelante, y desaparece la sensibilidad dolorosa y la térmica, conservándose la
táctil porque no sigue exactamente el trayecto de las otras. La lepra da un tipo de disociación
de la sensibilidad análoga a la de la siringomielia y por ello estos pacientes se queman sin
darse cuenta, o tienen úlceras indoloras.
El tálamo es un importante núcleo de células nerviosas, situado en la base del cerebro, al cual
confluyen todas las vías sensitivas mencionadas, y también las de los demás órganos de los
sentidos con excepción del olfato. Del tálamo parten nuevos contingentes de fibras que van a
terminar en distintos puntos de la corteza cerebral, las visuales en la corteza occipital, las
auditivos en la temporal, y las de la sensibilidad general en la parietal, en la parte lateral del
cerebro.
La corteza parietal ascendente de ambos lados es, pues, la estación terminal de las fibras de la
sensibilidad superficial y profunda. A su nivel hay una localización particular de las fibras que
le llegan, de modo que las que vienen de los miembros inferiores están en la parte alta y las de
los superiores en la baja, dibujando en forma invertida la imagen del cuerpo.
Localización de la sensibilidad en la corteza parietal del cerebro. A la derecha, en sombreado,
topografía de la zona sensitiva; a la izquierda, con aumento, detalle de la misma.